Bakulovirukset ovat hyvin laajalti levinnyt virusryhmä, joka infektoi lajispesifisesti niveljalkaisia (hyönteiset, hämähäkit ja äyriäiset). Viruksia tunnetaan yli 500 erilaista kantaa, joista useimmin käytetty ja bakulovirusten mallikappale on Autographa californica multicapsid nucleopolyhedro virus (AcMNPV).
1900-luvun puolivälissä bakulovirusten hyönteisiä infektoivat ominaisuudet huomattiin ja niiden käyttö tehokkaana biopestisidinä aloitettiin 1975. Käyttö sai myös aikaan runsaasti tutkimusta, jonka myötä selvitettiin bakulovirusten molekylaarista biologiaa, patologiaa ja ekologiaa. Bakulovirusten käytön turvallisuutta ja patogeenisuutta selkärankaisiin tutkittiin myös laajalti tuohon aikaan. Yhteenvetona on todettu, että erilaisissa turvallisuustesteissä ei havaittu virusten kykenevän infektioon tai aiheuttamaan allergisia reaktioita selkärankaisissa, eikä niissä myöskään todettu vasta-aineiden muodostumista koe-eläimille tai ihmisille ruoansulatuskanavan, hengitysteiden tai ihon kautta tapahtuvien altistumisten jälkeen.
Ensimmäinen bakuloviruksiin pohjautuva ekspressiovektori, jolla kyettiin tuottamaan proteiineja hyönteissoluissa esiteltiin vuonna 1983. Silloin käytetyimpiä solutyyppejä olivat Spodoptera frugiperda –peräiset SF-21 ja SF-9. Uudempana solutyyppinä ovat tulleet käyttöön Trichoplusia ni -peräiset High Five -solut. Bakulovirusten käytön proteiiniekspressioon tekee oivalliseksi se, että hyönteissolut ovat aitotumallisia (eukaryootti) soluja, ja siten kykenevät suorittamaan useille aitotumallisten solujen proteiineille tärkeät modifikaatiot paremmin kuin paljon käytetyt alkeistumalliset (prokaryootti) bakteerisolut. Esimerkkeinä modifikaatioista ovat mm. signaalipeptidin poistaminen, N-glykosylaatio, asylaatio, fosforylaatio ja karboksimetylaatio. Bakuloviruksia on alettu käyttää tästä syystä myös peptidinäyttötekniikan sovellutuksissa, joissa esiteltävän proteiinin post-translationaaliset modifikaatiot ovat oleellisia selektion kannalta.
1980-luvulla havaittiin, että bakulovirukset kykenevät penetroitumaan useisiin nisäkässolulinjoihin, mutta eivät kyenneet saamaan aikaan infektiota. Vuonna 1995 esiteltiin ensimmäisen kerran havainto, että muokatut AcMNPV:t kykenivät toimittamaan erillistä geenimateriaalia nisäkässoluihin. 1998 suoritettiin ensimmäinen ex vivo geeninsiirto hepatosyytti-solulinjaan ja vuonna 2000 julkaistiin tulokset joissa bakulovirusten todettiin kykenevän myös siirtämään geneettistä materiaalia in vivo adenoviruksiin verrattavalla tehokkuudella. Näiden havaintojen myötä tutkimus bakulovirusten käyttämiseksi geeniterapia-vektorina on kiihtynyt. Bakuloviruksilla on monia hyviä puolia perinteisesti käytettyihin geeniterapiavektoreihin verrattuna: kykenemättömyys jakautua nisäkässoluissa, suuri DNA-kapasiteetti, helppo valmistus ja alhainen sytotoksisuus. Huonoina puolina voidaan mainita mm. veren komplementin bakuloviruksia inaktivoiva vaikutus sekä ohimenevä (eng. transient) ekspressio. 2000-luvun puolivälin jälkeen virusten tutkiminen geeni- ja soluterapiakäyttöön on lisääntynyt. Uusimpina modifikaatioina ovat mm. lentiviruksiin siirretty gp64 pintaproteiini ja avidiini-gp64 fuusioproteiinilla päällystetyt bakulovirukset, joita on yhdessä raudasta koostuvien nanopartikkeleiden kanssa kyetty seuramaan MRIssä ensimmäisenä viruksena maailmassa.
Bakulovirukset koostuvat kahdesta suvusta, Nuclear polyhedrosis- viruksista (NPV) ja Granulosis- viruksista (GV). NPV:t tuottavat hyönteissoluissa infektion myötä suuria polyhedriini-proteiinimatrikseja, joiden sisällä on useita viruksia ja GV:t partikkeleita, joiden sisällä on vain yksi virioni.
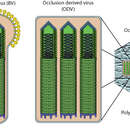
Bakulovirukset ovat hyönteissoluja infektoivia viruksia ja varsin lajispesifisiä. AcMNPV infektoi spesifisesti yli 30:tä lepidoptera –suvun hyönteislajia. Tämä suuri isäntälajien määrä on kuitenkin poikkeuksellista bakulovirusten keskuudessa. Infektoidessaan hyönteissoluja bakulovirukset esiintyvät kahdessa erilaisessa muodossa: niin sanottuna okluusioperäisenä viruksena (ODV: occlusion derived virus, joskus myös nimellä PDV eli polyhedron derived virus), joka sisältää useita viruskapsideita polyhedriinimatriksiin ympäröitynä, joka taas on päällystynyt vaipalla. Toinen muoto on silmikoituva virus (budded virus, lyhyemmin BV, joskus myös extra-cellular virion eli ECV, ekstrasellulaarinen virus), jolloin yksittäistä viruskapsidia ympäröi oma vaippa proteiineineen. Edellä mainittu ODV-muoto on tarpeen suojaamaan fyysisesti viruksia luonnossa, esim. puiden lehdillä. ODV-muodon polyhedriinimatriksi suojaa viruksia hyvin tehokkaasti ympäristön vaikutuksilta ja lisää siten virusten infektiivisyyttä ekosysteemissä. Joutuessaan hyönteisen ruuansulatuksen alkaaliseen ympäristöön polyhedriinimatriksi hajoaa vapauttaen ODV-virionit. Nämä ensimmäisen aallon virionit ovat hyvin tehokkaita tartuttamaan suoliston epiteelisoluja, mutta ne eivät ole juurikaan tehokkaita levittämään infektiota ympärilleen.
Silmikoituva virus -tyyppi on taas tarpeen kun primäärinen infektio hyönteisen ruuansulatuselimistön soluihin on saavutettu. BV-virukset ovat erikoistuneet solusta-soluun infektioon, jonka tarkoituksena on levittää systeemistä infektiota ympäri hyönteisen elimistöä. Solujen lyysautumisen alkaminen johtaa ODV- muodon tuotannon käynnistymiseen infektion myöhäisessä vaiheessa ja pian isännän kuolemaan. Polyhedriinin suojaamat ODV- virukset jatkavat infektion levittämistä ekosysteemissä. Kahden erilaisen muodon käyttö viruksen lisääntymisessä lienee syy sen hyvään menestykseen luonnossa.
Bakuloviruksen soluunotto on tärkeä vaihe niin sen luontaisen kiertokulun kuin erilaisten geeniterapian sovellustenkin takia. Soluunoton kaikkia vaiheita ei toistaiseksi tunneta, eikä myöskään olla täysin selvillä siitä, miten viruksen kapsidi päätyy solun tumaan. Soluunotto hyönteisissä on kuitenkin analoginen mekanismeiltaan nisäkässoluissa tapahtuvaan soluunottoon. Seuraavat mekanismit ovat siis tämänhetkisen tiedon mukaan yhteisiä AcMNPV:n tunkeutumisessa sekä hyönteissoluihin että nisäkässoluihin: tunkeutuminen soluun alkaa sitoutumisella isäntäsolun toistaiseksi tunnistamattomaan reseptoriin plasmamembraanilla, reseptorin kuroutumisella sisäänpäin ja endosyyttisen vesikkelin muodostumisella. Endosomin happamoituminen saa aikaan viruksen vaipan ja endosomin membraanin fuusion, joka vapauttaa nukleokapsidin sytoplasmaan. Viruksen kapsidi kuljetetaan tämän jälkeen tumaan replikaatiota varten.
Silmikoituva virus (BV) on rakenteeltaan sauvamainen ja sitä ympäröi membraani, jonka toisessa päässä on gp64–vaippaproteiineja peplomeerimäisenä rakenteena. Membraanin sisältä löytyy viruksen kapsidi, jonka sisällä viruksen sirkulaarinen genomi on. Okluusioperäinen virus (ODV) sisältää puolestaan useita kapsideita, ilman gp64-proteiinia, yhteisessä polyhedriiniproteiinista muodostuvassa matriisissa. AcMNPV:n genomi on 133 894 emäsparin mittainen ja muodostuu kaksisäikeisestä DNA:sta. Se koodaa potentiaalisesti 150 proteiinia.
Polyhedriini ja granuliini-proteiini (AcMNPV:n orf8 eli open reading frame 8, luettava geeni 8) ovat 245 aminohapon muodostamia proteiineja. Polyhedriini on myös hyvin konservoitunut proteiini eri bakuloviruskantojen keskuudessa. AcMPV:n polyhedriinipromoottori polh on hyvin käytetty proteiiniekspressiossa. Tämä johtuu siitä, että jopa puolet solun proteiineista voi infektion aikana muodostua tämän promoottorin alla. Polyhedriini ei ole oleellinen viruksen replikaatiolle tai sen infektiivisyydelle.
ODV-muotoisen viruksen polyhedrarakennetta ympäröi vaippa (eng. polyhedron envelope eli PE tai polyhedron calyx). Tämän vaipan pääasiallinen rakenneproteiini on PEP (Polyhedron envelope protein, pp34, orf131). Polyhedra-rakenteen toinen oleellinen proteiini on p10 (orf 137), jonka ekspressio tapahtuu myöhemmin infektion aikana. Myös se on oleellinen polyhedran stabiiliuden ja rakenteen kannalta.
BV:llä ja ODV:llä on hyvin samankaltaiset kapsidit, vain proteiini ODV-EC27:n on osoitettu löytyvän yksinomaan okluusiomuodosta. AcMNPV:n proteiini p6,9 (orf100) on DNA:han sitoutuva proteiini, joka on osallisena viruksen DNA:n pakkautumisessa sen kapsidin sisään. Se toimii spermassa histoneita korvaavien protamiinien tavoin ja on aminohappokoostumukseltaan samankaltainen. Se on oleellinen viruksen DNA:n pakkautumisessa kapsidin sisälle.
Mainitsemisen arvoinen kapsidiproteiini AcMNPV:lle on myös vp-39 (orf89), jota löytyy satunnaisesti kautta nukleokapsidin. Tämä viittaisi siihen, että se on kapsidin rakennekomponentti, erikoistuneen rakenteen sijaan. Proteiinin kysteiinitähteet ovat konservoituneet eri viruslajien keskuudessa, joka viittaisi niiden olevan oleellisia rakenteen kannalta.
Kapsidin päädyssä oleva ORF1629 (orf9) on erikoistunut kuppimainen rakenne, joka sulkee viruksen kapsidin. Viruksen kapsidin orientaatio infektion ja soluuntunkeutumisen aikana vaikuttaa kytkeytyvän ORF1629:n kautta kapsidin sisällä olevan DNA:n vapautumiseen kapsidista.
Bakuloviruksia ympäröivä vaippa koostuu kaksoiskerroksellisesta lipidikalvosta ja siihen liittyneistä proteiineista. Olennainen osa bakulovirusten rakennetta ovat niiden vaipan proteiinit, joilla on erilaisia rakennetta stabiloivia tai infektion kannalta olennaisia tehtäviä. ODV -ja BV -muotojen välillä on havaittavissa selviä eroja vaipan koostumuksen suhteen; ODV:n vaippa sisältää enemmän tyydyttyneitä rasvahappoja, kolesterolia ja tiheämmin proteiineja kuin BV:n vaippa. ODV:n vaippa koostuu tumakalvosta, kun taas BV:llä vaippa on peräisin solumembraanista. Tämän lisäksi osa eroista johtunee viruksen vaipan proteiinien osallistumisesta selektiivisesti membraanin ottamiseen isäntäsolun ulko- tai sisäpinnalta. BV-muotoinen virus, joka levittää infektiota systeemisesti, tarvitsee liikkuvamman vaipan ja erilaisen proteiinikoostumuksen kuin ODV, jonka vaipan proteiinit osallistuvat polyhedriinimatriksin kristallisoitumiseen.
Viruksen membraaniin on assosioitunut useita erilaisia proteiineja, niistä ODV-E66:n (orf46) tarkoituksena vaikuttaa olevan proteiinien targetointi, säilyttäminen ja/tai siirto ODV-muodon vaipalle. Samantapainen rakenne ja tehtävä vaikuttaa olevan myös ODV-E25:llä (orf94). p74 (orf138) -proteiinia ekspressoidaan infektion loppuvaiheessa ja geenin deleetio aiheuttaa virukselle kykenemättömyyden infektoida hyönteisiä ruuansulatuskanavan kautta.
Gp64 (orf128) on ainoa proteiini, joka löytyy yksinomaan BV-muodosta. Proteiini on lukukehyksen ennusteen mukaan 59 kDa:n kokoinen, mutta valmis proteiini on vahvasti glykolysoitu ja asyloitu ollen kooltaan 64 kDa. Geeniä säädellään aikaisella ja myöhäisellä (eng. late ja early) promoottorilla ja sen tuottama gp-64 proteiini on tärkein pohja fuusioproteiineille peptidinäyttötekniikassa. Gp64 on rakenteeltaan ja toiminnaltaan parhaiten tutkittu proteiini bakuloviruksissa, johtuen sen sopivuudesta erilaisiin tuotanto- ja näyttömenetelmiin.
Immunologisten ja biokemiallisten tutkimusten mukaan gp64 kerääntyy plasmamembraanille infektion alku- ja loppuvaiheissa. Viruksen nukleokapsidit tulevat vaipallisiksi työntyessään ulos solusta, jolloin viruksen vaippa muodostuu gp64-modifioidusta solumembraanista ja gp64-proteiineista. Itse viruksessa proteiinit ovat konsentroituneet viruspartikkelin toiseen päähän peplomeeri-rakenteiksi ollen olennaisia viruksen solusta-soluun infektiossa.
Gp64-proteiini sisältää N-terminaalisen signaalipeptidin ja C-terminaalisen ankkuridomeenin. Baculoviridae-heimon keskuudessa proteiini on erittäin konservoitunut. Runsaan glykolysaation lisäksi proteiini on asyloity, merkittävimmin palmitiinihapolla. Valmis gp64 trimerisoituu nopeasti disulfidisidosten kautta ja on membraanilla trimeerisessä muodossa. Trimerisaatiossa on osallisena proteiinin leusiini-vetoketju (amfipaattinen alfa-heliksi) rakenne.
Bakulovirukset ovat hyvin laajalti levinnyt virusryhmä, joka infektoi lajispesifisesti niveljalkaisia (hyönteiset, hämähäkit ja äyriäiset). Viruksia tunnetaan yli 500 erilaista kantaa, joista useimmin käytetty ja bakulovirusten mallikappale on Autographa californica multicapsid nucleopolyhedro virus (AcMNPV).
1900-luvun puolivälissä bakulovirusten hyönteisiä infektoivat ominaisuudet huomattiin ja niiden käyttö tehokkaana biopestisidinä aloitettiin 1975. Käyttö sai myös aikaan runsaasti tutkimusta, jonka myötä selvitettiin bakulovirusten molekylaarista biologiaa, patologiaa ja ekologiaa. Bakulovirusten käytön turvallisuutta ja patogeenisuutta selkärankaisiin tutkittiin myös laajalti tuohon aikaan. Yhteenvetona on todettu, että erilaisissa turvallisuustesteissä ei havaittu virusten kykenevän infektioon tai aiheuttamaan allergisia reaktioita selkärankaisissa, eikä niissä myöskään todettu vasta-aineiden muodostumista koe-eläimille tai ihmisille ruoansulatuskanavan, hengitysteiden tai ihon kautta tapahtuvien altistumisten jälkeen.
Ensimmäinen bakuloviruksiin pohjautuva ekspressiovektori, jolla kyettiin tuottamaan proteiineja hyönteissoluissa esiteltiin vuonna 1983. Silloin käytetyimpiä solutyyppejä olivat Spodoptera frugiperda –peräiset SF-21 ja SF-9. Uudempana solutyyppinä ovat tulleet käyttöön Trichoplusia ni -peräiset High Five -solut. Bakulovirusten käytön proteiiniekspressioon tekee oivalliseksi se, että hyönteissolut ovat aitotumallisia (eukaryootti) soluja, ja siten kykenevät suorittamaan useille aitotumallisten solujen proteiineille tärkeät modifikaatiot paremmin kuin paljon käytetyt alkeistumalliset (prokaryootti) bakteerisolut. Esimerkkeinä modifikaatioista ovat mm. signaalipeptidin poistaminen, N-glykosylaatio, asylaatio, fosforylaatio ja karboksimetylaatio. Bakuloviruksia on alettu käyttää tästä syystä myös peptidinäyttötekniikan sovellutuksissa, joissa esiteltävän proteiinin post-translationaaliset modifikaatiot ovat oleellisia selektion kannalta.
1980-luvulla havaittiin, että bakulovirukset kykenevät penetroitumaan useisiin nisäkässolulinjoihin, mutta eivät kyenneet saamaan aikaan infektiota. Vuonna 1995 esiteltiin ensimmäisen kerran havainto, että muokatut AcMNPV:t kykenivät toimittamaan erillistä geenimateriaalia nisäkässoluihin. 1998 suoritettiin ensimmäinen ex vivo geeninsiirto hepatosyytti-solulinjaan ja vuonna 2000 julkaistiin tulokset joissa bakulovirusten todettiin kykenevän myös siirtämään geneettistä materiaalia in vivo adenoviruksiin verrattavalla tehokkuudella. Näiden havaintojen myötä tutkimus bakulovirusten käyttämiseksi geeniterapia-vektorina on kiihtynyt. Bakuloviruksilla on monia hyviä puolia perinteisesti käytettyihin geeniterapiavektoreihin verrattuna: kykenemättömyys jakautua nisäkässoluissa, suuri DNA-kapasiteetti, helppo valmistus ja alhainen sytotoksisuus. Huonoina puolina voidaan mainita mm. veren komplementin bakuloviruksia inaktivoiva vaikutus sekä ohimenevä (eng. transient) ekspressio. 2000-luvun puolivälin jälkeen virusten tutkiminen geeni- ja soluterapiakäyttöön on lisääntynyt. Uusimpina modifikaatioina ovat mm. lentiviruksiin siirretty gp64 pintaproteiini ja avidiini-gp64 fuusioproteiinilla päällystetyt bakulovirukset, joita on yhdessä raudasta koostuvien nanopartikkeleiden kanssa kyetty seuramaan MRIssä ensimmäisenä viruksena maailmassa.