Prymnesium parvum ist eine Art (Spezies) von einzelligen Mikroalgen (etwa 10 µm lang und 3 bis 4 µm breit) aus dem Taxon der Haptophyta, dessen Mitglieder meist in mariner und eher tropischer Umgebung leben.
P. parvum ist biflagellat (doppelt begeißelt), was es ihr ermöglicht, sich zu bewegen. Man findet sie daher auch in der Regel schwebend in der Wassersäule, am häufigsten in Ästuar- oder Meeresgewässern. Sie kann aber auch in Binnengewässern vorkommen, die einen relativ hohen Mineralgehalt aufweisen. Die Typlokalität ist ein Brackwasserteich auf der Isle of Wight, England.[1][2]
P. parvum kommt heute kosmopolitisch (weltweit) vor, unter anderem in der Ostsee, Europa, China, Australien, den USA und Marokko. P. parvum kann sie schädliche Algenblüten (englisch harmful blooms, HABs) in Binnen- und Küstengewässern hervorrufen, die zu verheerenden Fischsterben mit ökologischen sowie wirtschaftlichen Schäden führen.[1][2]
Wegen der Pigmente, die ihr einen goldgelben Schimmer verleihen, wird P. parvum manchmal als "Goldalge" (en. golden algae, golden brown algae) genannt. Die Bezeichnung Goldalgen ist jedoch ein nicht-taxonomischer Begriff, mit dem verschiedene Gruppen der Stramenopilen aufgrund ihrer Farbe subsumiert werden, darunter die Goldbraunen Algen (Chrysophyceae), die Goldgrünen Algen (Bacillariophyta) oder die Kieselalgen (Bacillariophyta). Um Verwechslungen zu vermeiden sollte diese Bezeichnung daher in wissenschaftlichen und juristischen Texten vermieden werden.
Die Spezies Prymnesium parvum wird in Unterarten oder Formen wie folgt untergliedert:[3][4][5][6][7]
Zu den Formen sind jeweils eine Reihe von Stämmen bzw. Isolaten bekannt.
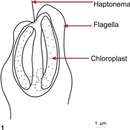
P. parvum ist nur ca. 10 µm groß und kann vermutlich vier morphologisch unterschiedliche Formen annehmen. Zwei dieser Stadien sind biflagellate (doppelt begeißelte) haploide Zelltypen. Ein weiteres mögliches Stadium ist ein biflagellater diploider Zelltyp. Neben diesen motilen (beweglichen) Formen gibt es eine unbewegliche Form ohne Geißeln, was ein Ruhestadium sein könnte. Die begeißelten Formen haben außer den beiden Geißeln eine Haptonema, d. h. eine spezielle äußere nadelartige Struktur, die das Anhaften an Oberflächen ermöglicht. Es werden zwei Unterarten unterschieden:[1]
Die Zellen von P. parvum haben zwei sattelförmige Chloroplasten, die in der Regel gelbgrün bis olivgrün gefärbt sind. Die Geißeln sind zwischen 12 und 15 µm lang, die flexible, nicht gewundene Haptonema zwischen 3 und 5 µm. Jede Zelle hat zwei Schichten von Körperschuppen unterschiedlichen Typs, die Schuppen der äußeren Schicht haben schmale, gebogene Ränder, die der inneren Schicht breite, stark gebogene Ränder.[1]
P. parvum lebt weder rein autotroph (etwa von der Photosynthese) noch rein heterotroph (räuberisch) lebend.
Das Verhalten dieser Alge ist ungefähr wie bei der (nicht-taxonomischen) Gruppe der Fleischfressenden Pflanzen: Mit seinem Chloroplasten kann P. parvum im Prinzip Photosynthese (Photoassimilation) betreiben. Bei Nährstoffmangel (insbesondere bei Phosphatmangel) oder im Schwarm kann P. parvum andere Organismen „fressen“ (Phagozytose). Der Stoffwechsel dieses Mikroorganismus wechselt unter diesen Umständen in den heterotrophen Modus, und der Mikroorganismus wird zum „Räuber“, indem er Beute oder tote Zellen phagozytiert. Dies ermöglicht der Mikroalge ein Leben im Dunkeln, etwa durch Abweiden von bakteriellen Biofilmen, Möglicherweise befriedigt P. parvum dann seinen Bedarf an Phosphaten durch den Verzehr von Bakterien.[11][12][13]
P. parvum kann eine breite Palette von Stickstoffquellen nutzen, einschließlich Ammonium, Nitrat, Aminosäuren (mit einer gewissen pH-Abhängigkeit), Kreatin, ist aber nicht in der Lage, Harnstoff zu nutzen.[13]
P. parvum produziert Dimethylsulfoniopropionat (DMSP) und andere Polyole, deren Funktionen noch unbekannt sind, die aber mit Anpassungen (Osmoregulation) an ungewöhnlich salzhaltige oder mineralisierte Umgebungen verbunden sein könnten.[14][13]
Unter bestimmten Konkurrenzbedingungen reduziert P. parvum seine Aktivität geht in ein Ruhestadium („Zyste“, en. “cyst”, dormant/resting stage) über (im modernen Sprachgebrauch bezeichnet bei Mikroorganismen der Begriff Zyste eine Dauerform ohne Stoffwechsel, insbesondere auch ohne Photosynthese; andernfalls liegt lediglich ein unbegeißeltes, icht-motiles Stadium vor – was im früheren Sprachgebrauch nicht unterschieden wurde).[13][2] In der neueren Literatur ist beispielsweise die Rede von „stationären Wachstums-/Todphasen“ (en. stationary growth/death phases).[15] Auf die produzierten Toxine (Prymnesine) wird weiter unten noch gesondert eingegangen.
P. parvum wächst in einem Salzgehalt (Salinität) im Bereich von 0,5 - 30 psu (Practical Salinity Unit) mit einem Optimum bei 15 psu. Allerdings scheinen Stämme, die an verschiedenen Orten gesammelt wurden, unterschiedliche Salzgehaltstoleranzen zu haben. Ein Stamm namens LB 2797 (isoliert aus dem Colorado River in Texas) zeigt ein biphasisches Wachstumsmuster, d. h. die maximalen Zelldichten nahmen mit steigendem Salzgehalt von 5 bis 15 psu zu, nahmen aber bei höheren Werten in der Laborkultur wieder ab.[16]
Während Blüten von P. parvum in der östlichen Hemisphäre bereits seit den frühen 1900er Jahren dokumentiert sind, hat sich die Art seitdem weit verbreitet. Es wurden Blüten überall im Süden der USA, sowie in einigen nördlichen Regionen beobachtet.[1][2]
Diese in Flussmündungen sehr häufige Art lebt normalerweise nur im Brackwasser, wird aber zunehmend auch im Süßwasser gefunden, vor allem in den Vereinigten Staaten (z. B. in Texas seit 1985).[17] Dies gilt als besorgniserregend gilt, insbesondere bei Fischern, die in bestimmten Seen und Flüssen bereits Fischsterben beobachtet haben.[18] Sie kommt in der Natur in hellen, offenen Umgebungen vor, aber im Labor kann ihr Wachstum durch zu viel Licht gehemmt werden (Photoinhibition)[19].
In Florida wurde in den 2000er Jahren auch beobachtet, dass Fische durch Blüten dieser Alge in Hinterhof- und Golfplatzteichen, oft in Küstenregionen, getötet wurden.[2]
Algenblüten treten in der Regel vom Spätwinter bis zum Sommer auf. Sie führen dazu, dass sich das Wasser grün bis gelblich verfärbt und sich Schaum bildet, wenn das Wasser aufgewühlt wird (an Wehren, Dämmen, Ufern).[2] Die Algenblüten bilden sich besonders leicht im Brackwasserbereich, da die von P. parvum gebildeten Toxine dort leichter kritische Konzentrationen erreichen können.[20]
Im letzten Jahrhundert haben zunehmend saisonale Fischsterben in Verbindung mit den toxischen Algenblüten von P. parvum weltweit Aquakulturen und einheimische Fisch-, Schalentier- und Molluskenpopulationen zerstört. Langanhaltende Blüten von P. parvum können zu großen Störungen der lokalen Ökologie und hohen finanziellen Verlusten führen.[21][20]
Die toxische Wirkung von P. parvum u. a für Fische wird auf eine Gruppe organo-chemischer Verbindungen zurückgeführt, die Prymnesine genannt werden und chemisch Polyether mit einer großen Leiterrahmenstruktur (en. supersized ladder-frame polyether compounds) sind.[15] Es sind biochemisch komplexe und hochmolekulare Verbindungen.[22] Diese Prymnesine werden derzeit (Stand 2019) in drei Typen A, B und C unterteilt (9 vom A-Typ, 12 vom B-Typ und 30 vom C-Type).[23] Es ist gut möglich, dass noch nicht alle von dieser Alge abgesonderten Toxine identifiziert wurden.
Sie zeigen eine starke zytotoxische, hämolytische, neurotoxische und ichthyotoxische Wirkungen aufweisen.[21][15][22]
Bei Stress sondert die P. parvum diese chemischen Verbindungen in das Wasser ab. In Verbindung mit im Wasser befindlichen Kationen (z. B. Magnesium-Ionen Mg++ oder Calcium-Ionen Ca++) bilden sich dann die effektiven Toxine. Diese sind also abhängig von der Chemie des Wassers, wobei meist eine Kombination verschiedener Toxine vorhanden ist. Die genaue Zusammensetzung, d. h. der Anteil eines jeden der verschiedenen Prymnesine hängt vom jeweiligen Stamm von P. parvum ab.[15][23] Der Stamm K-0081 enthielt ~5 mal mehr Toxin als der Stamm K-0374.[15]
Diese Toxine könnten als Gifte wirken, die es diesen Algen normalerweise ermöglichen, andere einzellige Organismen zu fangen und fressen (phagocytieren). Man vermutet, dass es ihre Funktion ist, Beutetiere zu verlangsamen oder unbeweglich zu machen, damit sie dann zur Beute der Algen werden.
Werden die Toxine aber von einer großen Menge Algen massiv ins Wasser abgegeben bzw. aus ihren Vorstufen erzeugt, dann wirken sie auch auf höhere tierische Organismen wie Fische. Angegriffen werden zuerst schlecht geschützte und exponierte Zellen, z. B. auf der Oberfläche der Kiemen von wirbellosen Wassertieren und von Fischen, sowie auf deren Flossen. Sie verhindern die ordnungsgemäße Funktion der Wasserregulierung in der Zelle (Osmoregulation), die durch Vergiftung und/oder Wasserüberschuss abstirbt. Nachdem die erste Zellschicht zerstört ist, werden die nächsten Schichten angegriffen. Wenn ein Blutgefäß betroffen ist, kommt es zur Blutung. Durch die erodierten Kiemen gelangen die Toxine in das Blut- und Kreislaufsystem des Fisches. In der Folge werden auch die inneren Organe geschädigt. Die ersten sichtbaren Symptome sind, dass der Fisch sich wie bei Sauerstoffmangel im Wasser verhält; er pendelt zwischen der Oberfläche, wo er nach Luft zu schnappen versucht, und dem Grund, wo er ruht und stirbt.[24]
Neuere Erkenntnisse zeigen, dass diese Art nur unter Umweltstress Toxine produziert, z. B. bei einem Überangebot an planktischen Räubern (Zooplankton). Dinoflagellaten der Spezies Oxyrrhis marina (Oxyrrhinales). Offenbar benutzt P. parvum die Toxine in diesem Fall zur Abwehr der eigenen Fressfeinde. Jedenfalls nahmen die Dinoflagellaten weniger P. parvum als Nahrung auf als andere marine Algen, etwa Cryptophyceen der Gattung Rhodomonas. Die Toxine bewirken offenbar eine Verlangsamung ihrer Aufnahme- und Verdauungskapazität der Dinoflagellaten.[25]
In ähnlicher Weise kann der Stress durch einen Mangel an Stickstoff oder Phosphor die Produktion von Toxinen auslösen. Vermutlich profitiert P. parvum dann nicht nur durch leichteres Beute machen, sondern auch von den Spurenelementen profitieren die von den abgestorbenen Individuen der anderer Arten freigesetzt werden – dies könnte ein Weg sein, um in einer Umgebung konkurrenzfähiger zu sein.
Prymnesin-1 (Prym1) ist eine zu den Typ-A-Prymnesinen gehörende Chemikalie. Dies ist eine Klasse von P. parvum gebildeter hämolytischer Phykotoxine (Algentoxine), Polyether mit Leiterrahmen-Aufbau.[26][27][28]
Der Stoff ist bekannterweise für Fische giftig ist und fühtz weltweit immer wieder zu massenhaften Fischsterben führt, unter anderem in Texas und England.
Prymnesin-2 (Prym2) ist eine weitere organische Verbindung aus der Klasse der Typ-A-Prymnesine, die von P. parvum gebildet wird.[26] Auch diese Substanz hat starke hämolytische und ichthyotoxische Eigenschaften. In gereinigter Form erscheint sie als blassgelber Feststoff.[20]
Prymnesin-B1 (PrymB1) ist eine weitere Chemikalie aus der Klasse der Prymnesine, die von P. parvum gebildet werden. Auch hier ist die Giftigkeit für Fische bekannt. Es handelt sich um ein Typ-B-Prymnesin, das sich im Vergleich zu Typ-A-Prymnesinen (wie Prymnesin-2) durch die Anzahl der Rückgratzyklen unterscheidet.[26]
Eine weitere Chemikalie aus der Klasse der Typ-B-Prymnesine ist Prymnesin-B2 (PrymB2).[15]
Die Typ-C-Prymnesine (PrymC) sind die Gruppe der Prymnesine mit der größten Diversität. Dieser Typ ist derzeit (2019) noch am wenigsten verstanden.[23]
Diese Toxine scheinen keine Auswirkungen auf Rinder zu haben, die mit diesem Toxin kontaminiertes Wasser trinken, oder auf Aasfresser, die durch die Toxine vergifteten Fische fressen. Das unterscheidet sie von anderen Algen, die sich auf die gleiche Weise vermehren können (bestimmte Cyanobakterien, Algen, die für „Rote Tiden“ verantwortlich sind etc., teilweise auch „Ultragifte“). Eine Hypothese, um dies zu erklären, ist, dass die Alge und ihre Toxine vom pH-Wert abhängig sind, und ein basisches oder zumindest nur wenig saures Milieu benötigen. Durch den Säuregehalt des Magens würden die Toxine zerstört werden. Darüber hinaus könnten die Haut und auch Schleimbildung bei Landwirbeltieren wie es die Säugetiere sind ebenfalls besser schützen als eine Fischhaut. Insgesamt sind die Auswirkungen auf den Menschen nicht gut untersucht. Nach Angaben des Texas Department of State Health Services wird aber auf jeden Fall empfohlen, keine Fische zu verzehren, die durch eine planktonischen Blüte von P. parvum in Mitleidenschaft gezogen wurden.[29]
P. parvum verursacht aufgrund der Toxizität der von ihr produzierten Moleküle (u. a. die Prymnesine) in Nordamerika seit Jahren ökologische und toxikologische (ökotoxikologische) sowie wirtschaftliche Probleme.[30]
Diese Toxine töten viele kaltblütige und aquatische sowie auch semiaquatische Wirbeltiere (z. B. Salamander) bereits bei niedrigen Dosen und beeinträchtigen den Rest der planktischen Gemeinschaft, so dass direkt oder indirekt das gesamte Ökosystem betroffen ist. Die Wirkung ist allelopathisch, d. h. die Alge hemmt das Wachstum von Cyanobakterien, Dinoflagellaten und tötet Wimpertierchen sowie Kieselalgen, wodurch sie (besser) gedeihen kann.[22]
Diese Alge kann auch in einigen küstennahen Roten Tiden (vgl. Heterosigma akashiwo) vorkommen, wo sie zusätzlich zum Fischsterben beitragen könnte.[31]
Die Erstbeschreibung von P. parvum stammt aus dem Jahr 1937, die Typlokalität ist jedoch ein Brackwasserteich auf der Isle of Wight, England, UK. In Nordamerika wurde die Spezies 1985 entdeckt. Es ist nicht bekannt, ob sie in Nordamerika bereits in vorkolumbiasnischer Zeit (vor der Ankunft der Menschen aus dem Westen) und vor der Entwicklung der Industrie dort bereits natürlicherweise existierte oder ob sie vom Menschen eingeführt wurde (wie bei einigen anderen Algenarten, die leicht invasiv werden können und auch in bestimmten Gegenden invasiv wurden, und die bei früheren Bestandsaufnahmen nie gefunden wurden). Im Süßwasser scheint sie durch saure und mineralisierte Einleitungen aus dem Bergbau oder von Anwohnern begünstigt zu werden. Beispielsweise wurden in der Umgebung des Lake Granbury (einem Stausee in Nordtexas in der Nähe von Granbury, Texas), der von Episoden ökotoxischer Algenblüten betroffen war, die Abwässer vieler Anwohner ohne Klärung direkt in den See eingeleitet.[32]
Die Mikroalge wurde in vitro oder in Bioreaktoren kultiviert, um sie zu studieren.[33] Wegen ihres hohen Lipidgehalts und der relativ einfachen Kultivierung geschah dies darüber hinaus auch im Rahmen von Biokraftstoff-Projekten.[34][35][7]
Die wirtschaftlichen Auswirkungen der durch P. parvum hervorgerufenen Algenblüten bzw. Fischsterben variieren je nach Größe des betroffenen Gewässers, seinem Fischreichtum und dem Ort der Ausbrüche. Es drohen Einnahmeverlusten für die Fischerei (Berufs-, aber auch Hobbyfischerei) oder für den sanften Tourismus. Insbesondere in Texas scheinen die wichtigsten direkten negativen wirtschaftlichen Auswirkungen Angelführer, Betreiber von Sommercamps, Parks, Hotels, Motels, Restaurants, Bekleidungsgeschäfte, Tankstellen usw. zu betreffen. Eine indirekte oder sekundäre Auswirkung ist die Verschlechterung des Images der betroffenen Gebietes, verbunden mit weiter verminderten Einnahmen. Eine wirtschaftliche Bewertung wurde für den Possum Kingdom Lake (PKL, ein Stausee am Brazos River, Texas) durchgeführt. Hier gab es Algenblüten durch P. parvum von Januar bis Juli 2001 und erneut im Jahr 2003. Die Algenblüte 2001 führte zu einem finanziellen Verlust von 2,8 Mio. US$ für die lokalen Gemeinden allein durch den Rückgang der Besucher.[36]
P. parvum wird parasitiert von Viren der vorgeschlagenen Spezies „Prymnesium parvum DNA virus BW1“ (PpDNAV, PpDNAV-BW1)[38] Dieses Riesenvirus wurde als Algenvirus zunächst mit der Virenfamilie Phcodnaviridae (Ordnung Algavirales) in Verbindung gebracht, scheint aber als mutmaßliches Mitglied einer Klade mit der vorläufigen Bezeichnung „OLPG“ (en. Organic Lake Phycodna (Virus) Group) eher den Mimiviridae (Ordnung Imitervirales) nahe zustehen oder anzugehören.[37][39][40]
Prymnesium parvum ist eine Art (Spezies) von einzelligen Mikroalgen (etwa 10 µm lang und 3 bis 4 µm breit) aus dem Taxon der Haptophyta, dessen Mitglieder meist in mariner und eher tropischer Umgebung leben.
P. parvum ist biflagellat (doppelt begeißelt), was es ihr ermöglicht, sich zu bewegen. Man findet sie daher auch in der Regel schwebend in der Wassersäule, am häufigsten in Ästuar- oder Meeresgewässern. Sie kann aber auch in Binnengewässern vorkommen, die einen relativ hohen Mineralgehalt aufweisen. Die Typlokalität ist ein Brackwasserteich auf der Isle of Wight, England.
P. parvum kommt heute kosmopolitisch (weltweit) vor, unter anderem in der Ostsee, Europa, China, Australien, den USA und Marokko. P. parvum kann sie schädliche Algenblüten (englisch harmful blooms, HABs) in Binnen- und Küstengewässern hervorrufen, die zu verheerenden Fischsterben mit ökologischen sowie wirtschaftlichen Schäden führen.
Wegen der Pigmente, die ihr einen goldgelben Schimmer verleihen, wird P. parvum manchmal als "Goldalge" (en. golden algae, golden brown algae) genannt. Die Bezeichnung Goldalgen ist jedoch ein nicht-taxonomischer Begriff, mit dem verschiedene Gruppen der Stramenopilen aufgrund ihrer Farbe subsumiert werden, darunter die Goldbraunen Algen (Chrysophyceae), die Goldgrünen Algen (Bacillariophyta) oder die Kieselalgen (Bacillariophyta). Um Verwechslungen zu vermeiden sollte diese Bezeichnung daher in wissenschaftlichen und juristischen Texten vermieden werden.









{kind=link}