Taxonomy and Phylogeny
In 1817, Nees first described Alternaria altenata (original name is Alternaria tenuis) and established the genus Alternaria, with Alternaria alternata as the type species (Rotem, 1994). Then the dispute about its taxonomy started. In 1925, Fries described the genus Macrosporium and replaced the genus Alternaria by Macrosporium. And then, he mentioned Alternaria under the genus Torula. Finally, Fries acknowledged the exit of Alternaria (Tweedy and Powell, 1963). So, Alternaria alternata has the following synonyms: Macrosporium fasciculatum Cooke & Ellis (1817), Torula alternata Fr. (1832), Alternaria fasciculata (Cooke & Ellis) L.R. Jones & Grout (1897), and Alternaria rugosa McAlpine (1896)(Wikipedia). In 1912, it was named Alternaria alternata (Fr.) Keissl. (1912)
Alternaria alternata belongs to the following group (Wikipedia):
Kingdom: Fungi
Phylum:Ascomycota
Class:Dothideomycetes
Subclass:Pleosporomycetidae
Order:Pleosporales
Family:Pleosporaceae
Genus:Alternaria
Species:Alternaria alternata
Alternaria alternata also belongs to Deutermycota, an artificial phylum and containing all the fungi whose sexual structure has not been observed or associated with its asexual structure.
Morphology
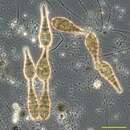
The genus Alternaria is characteristic by the large conidia. They are produced on chains, light brown to brown, ovoid or obclavate shaped, muriform and divided by transverse and vertical walls, with or without beak at the tip. The hyphae and conidiophore are light brown and septate. The dark pigment can prevent the fungal organism from damage caused by UV-radiation (Pusz, 2009).
The species can be divided into three groups according to the number of conidia on the conidiophore: Noncatenatae with single conidia; Brevicatenatae with short chains containing three to five conidia; Longicatenatae with long chains containing 10 spores or more, and Alternaria alternata belongs to this group (Rotem, 1994).
However, the microscopic differences between Alternaria species are not significant, and the character of every species varied depending on the conditions of growth. So, the identification of species is very difficult. Generally, the maximum width of spore is from 5-13 to 6-24 nm (the mean is 9 to 17 nm), and the length of spore is from 16-37 to 35-110 nm (the mean is 37 to 69 nm). The number of transverse septa is from 0 to 23. The beak length is from 4 to 90 nm. The conidiophore length is from 20 to 300 nm (Rotem, 1994). Usually, The conidia formed in natural habitats are larger, have longer beaked tip, and are more uniform than those formed in common agar media. And, under low temperature and dry conditions, larger conidia with shorter beak are produced (Misaghi et al, 1977). On the culture, it is a grey to brown, downy or cotton colony.
Ecology
Alternaria species is a hardy fungus and can live in extreme conditions. Alternaria alternata can pass the winter in the soil, seed, infected crop debris or perennial host tissue, such as bark, nodes, and scaly leaves as mycelia and or conidia. Some strains can produce survival structure, microsclerotia, to resistant the unfavorable condition.
While some Alternaria species need the stimuli to initiate the conidiophore formation and sporulation, Alternaria alternata can sporulate easily without triggering (Rotem, 1994). The spores of Alternaria alternata can be propelled into air by a shift from wetness to dryness, a rapid increase in humidity or exposure to the red light (Leach, 1975). Due to it big spores and dark pigment, Alternaria alternata can float in the air and avoid the damage from UV-radiation. So, airborne dispersal is its main dispersal method. So, it can be spread all over the world, such as North America, Africa, Europe, Asia, and Austria.
Alternaria alternata has a wide host range. Almost every host affected by Alternaria species is also affected by Alternaria alternata (Rotem, 1994). Alternaria alternata prefers to infect the senescent, heavily stressed or damaged plant tissue. On some plants, it is the secondary invader, appearing on the lesion caused by other Alternaria species (tamato). Alternaria alternata also can infect the intact and healthy plant tissue. Sometimes, it can penetrate into the flower, mycelia and conidia developing on seeds in the fruit (Halfon-Meiri and Rylski, 1983). The frequently reported plants can be infected by Alternaria alternata include: tobacco, citrus, pear, apple, wheat, tomato, bean, cotton, brassicaceae, pepper, cucurbits, potato, strawberry, and beet (Rotem, 1994).
The symptoms on the leaves are brown and circular lesions with sharply defined margin and yellow layers surrounded. Inside the lesions are many dark concentric circles like a target. With the development of disease, the lesions can develop and merge into many irregular areas. Under the favorable condition, the conidia can be seen on the lesion surface, giving the lesions a dusty appearance (Brown spot: Alternaria alternata (Fr.) Keissl).
Allergy
Allergic reactions caused by fungal mold have been recognized since 1726. In the past 300 years, it is reported that many fungi can cause the allergic reactions, such as Penicillium, Chaetomium, and wheat rust (Sanchez and Bush, 2001). Deuteromycetes has been recognize to contain most of the allergenic fungi; of which Alternaria alternata is one of the most important fungi that can cause asthma and rhinitis (Bush and Prochnau, 2004). Not only the intact spores can produce and transfer the allergens, the fragmented hyphae and spores also can transfer the allergens. And, they can cause more asthma than intact spores because they can respiratory tract (Pulimood et al, 2007). And, allergy content in the spores can vary with the developmental conditions (Green et al, 2003). In the United State, almost 3.6% of the population is sensitive to Alternaria alternata in the skin test, according to a survey covered a large-scale population (Gergen et al, 1987). In the outdoor environment, the disperse of Alternaria alternata spores occurs in the dry and windy day period, usually get the peak levels in the sunny afternoon. In North America, the dispersal of spore can get the peak level in late summer and early autumn. There is no detection of Alternaria alternata spores in winter with the snow cover and colder temperature (Bush and Prochnau, 2004). In the indoor environment, Alternaria alternata spores can be found everywhere, such as the kitchen, floor, ceiling, bed, and living room. So, the asthma caused by Alternaria alternata is a big problem to human.
Alternaria alternata is a fungus causing leaf spots, rots, and blights on many plant parts, and other diseases. It is an opportunistic pathogen on over 380 host species of plant.
It can also cause upper respiratory tract infections[1] and asthma in humans with compromised immunity.
Alternaria alternata has many different hosts depending on its forma specialis.
A. a. f. sp. lycopersici (AAL) infects only certain cultivars of tomato plants and is often referred to as Alternaria stem canker of tomato. AAL's main symptom is cankers in the stem. It resides in seeds and seedlings, and is often spread by spores as they become airborne and land on plants. It can also spread throughout other plants.[2] Under severe infection, lesions enlarge and become coalesced causing blighting of the leaves. This symptom progression occurred in research done in Pakistan: the symptoms on affected tomatoes started with yellowing and browning of the lower leaves, then began developing on the leaf tips and along the margins of the leaf petiole. This progression continued until the entire leaves were covered in diseased tissue and then fell off.[3] In addition to necrotic leaves and petioles, plants are found to have severe defoliation, with considerable yield losses when it occurs before flowering.[2] The tomato fruit can also be infected as well, with brown cankers dotting them and making them inedible. Once the disease has spread to a certain point, little can be done to save the tomato plant.
There are several host factors that affect disease development. For example, various signaling pathways in tomato plants affect their susceptibility to AAL.[4] Salicylic acid promotes resistance to AAL and antagonizes the ethylene response. Ethylene controls the synthesis of jasmonic acid, which is a necessary pathway for immunity. Independently of each other, salicylic acid, ethylene, and jasmonic acid can influence the susceptibility of tomato to AAL. Diagnosis of AAL is often from observing signs and symptoms from this fungal pathogen. In addition, a tomato cultivar's resistance to a toxin produced by AAL also affects disease development.
In order to survive, Alternaria alternata needs a moist warm environment. It is often found in areas with humid climates, or where there has been significant rainfall.[2] The fungus lives in seeds and seedlings, and is also spread by spores. This disease flourishes in dead plants that have been left in gardens over winter. Additionally, when dead infected debris is added to compost pile it can spread to other vegetables throughout the garden.
There are no insect vectors for this disease. This means that using insecticides has no effect on the spread of this pathogen. However, there are several cultural practices that can be followed to suppress this fungal pathogen's impact. The disease first occurs in the host's exposed leaves. Plants planted with rows in an east–west direction have more severe disease than do plants planted north–south. This implies that if one plants tomato plants in a north–south manner they will be less susceptible. It is also suggested to highly monitor plants in April through June. This is when the pathogen is most prevalent. If monitoring indicates the presence of AAL, it is suggested to begin late-spring treatments of fungicide about mid-April. However, if a garden has a history of disease, it is advised to take extra measures. This can be done by treating tomatoes in mid to late April and 2 to 3 weeks later by applying a fungicide.[5] Because this is a fungal pathogen that thrives off of wet environments, overhead irrigation is never advised when irrigating. This causes the moisture to remain on the leaf tissue and increase susceptibility to the disease and provides an optimal environment for the fungus to survive and grow. Furrow irrigation or drip irrigation systems allow the plant to remain dry.
Overall, AAL thrives in moist warm environments. Cultural practices for preventing this disease include planting tomatoes in a row north to south, monitoring plants heavily April through June, and using a drip irrigation system to keep as much plant tissue dry and free of favorable environments for this pathogen.
While γ-aminobutyric acid (GABA) has no direct fungicidal activity on A. alternata,[6] it does induce resistance in tomato (Solanum lycopersicum).[6] Some or all of that resistance is by activation of the tomato's own enzymes exerting an antioxidant effect.[6]
Teleomorph of Alternaria alternata is thought to be Clathrospora diplospora: this has yet to be confirmed.[7] If this is correct, as a result this pathogen would be propagating itself via asexual spores called conidia.[8] These conidia are produced in lesions on mature or dying leaves.[8] Their production can begin in as few as ten days after the first symptoms appear, and can continue for to up to fifty days.[9] A. alternata's conidia disperse via air currents, and their release from the lesions can be triggered by rainfall, or even just a sudden drop in humidity.[9] When the conidium lands on a leaf, it will wait until the nighttime dew, and then germinate.[8] It can either enter through the stomata, or penetrate directly through the top of the leaf, using its appressorium, infecting the leaf within 12 hours.[8]
At the cellular level, AAL produces toxins that are essential for pathogenicity on tomato.[10] This host specific mycotoxin is called fumonisin B1. It was identified and confirmed by research conducted on fast atom bombardment and ion spray mass spectrometry.[11] Thus, tomatoes that are resistant to this pathogen may be resistance to this specific toxin. Resistance to the pathogen in tomato is inherited as a single gene expressing complete dominance. However, sensitivity to the fumonisin B1 gene is controlled by a single locus with two alleles expressing incomplete dominance when heterozygous.[12] In addition to resistance to the specific gene, resistance can be found from signaling pathways.
At the organismal level, AAL grows very slowly. This makes it so its presence is often not known until seedlings become larger and are transplanted into the garden. A fungicide may be used to save the plants once they are infected; however, the disease cannot be completely eradicated.[5]
Ultimately, pathogenicity of this organism depends on a specific tomato cultivar's resistance to an AAL strain's specific fumonisin B1 variant. Additionally, the best way to prevent this pathogen from producing disease on tomato plants is to ensure the tomatoes are resistant cultivars.
Thought to be Clathrospora diplospora (to be confirmed).[7]
{{cite journal}}: CS1 maint: DOI inactive as of December 2022 (link) Alternaria alternata is a fungus causing leaf spots, rots, and blights on many plant parts, and other diseases. It is an opportunistic pathogen on over 380 host species of plant.
It can also cause upper respiratory tract infections and asthma in humans with compromised immunity.

