Kristofer M. Helgen, C. Miguel Pinto, Roland Kays, Lauren E. Helgen, Mirian T. N. Tsuchiya, Aleta Quinn, Don E. Wilson, Jesús E. Maldonado
Zookeys
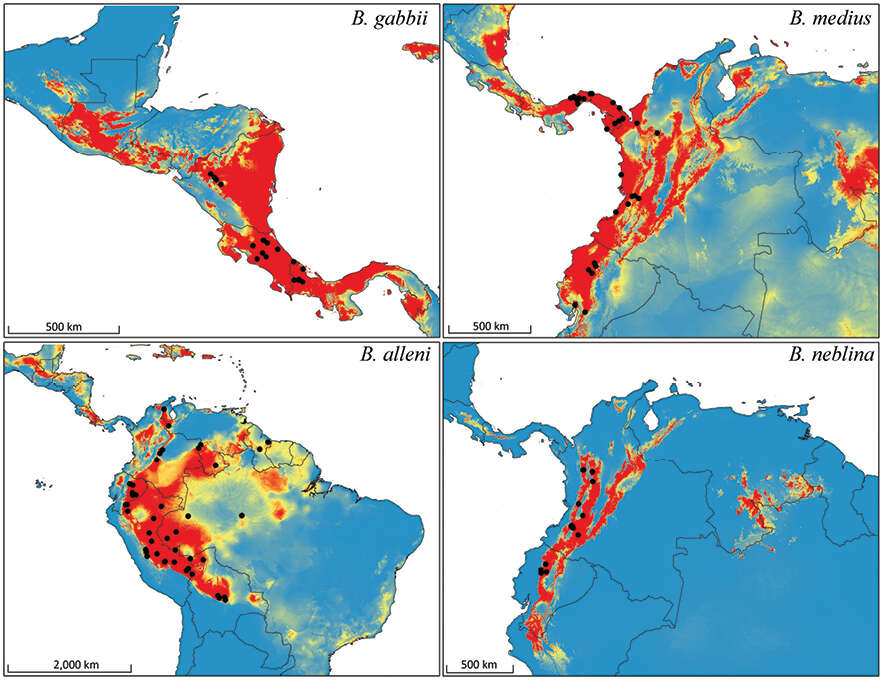
Figure 11.Bioclimatic distribution models and localities for Bassaricyon species. Models from MAXENT using all vouchered occurrence records, 19 bioclimatic variables, and one potential habitat variable.
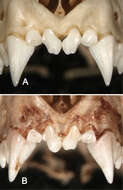
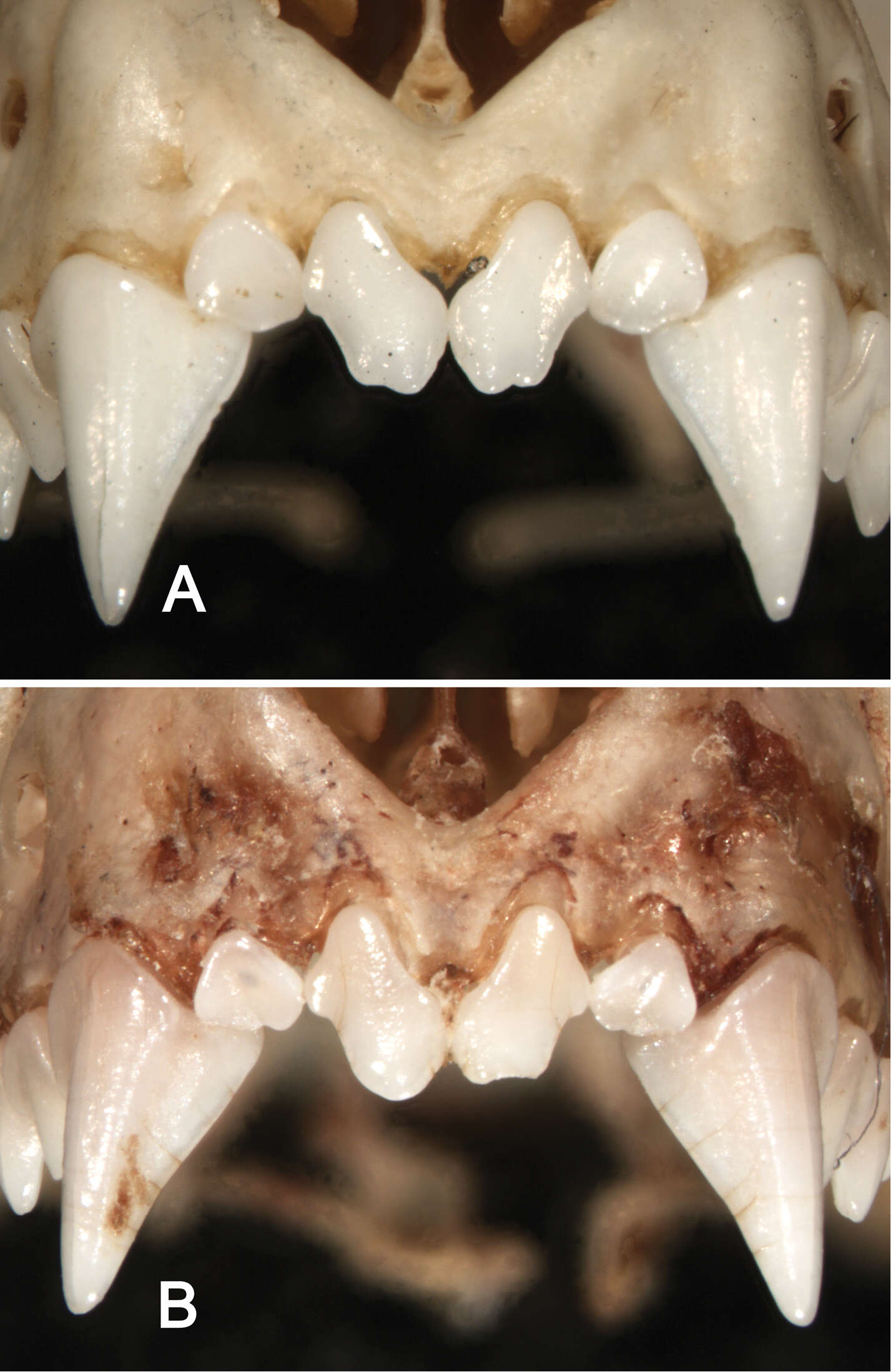
Figure 8.Anterior views of the upper incisors and canines in Sturnira bakeri (A, QCAZ 14635 ♀) and Sturnira burtonlimi (B, ROM 104294 ♂) illustrating taxonomic differences in the number of cuspids of the upper inner incisor (I1). In Sturnira bakeri the I1 is bicuspidate. In Sturnira burtonlimi, however, the I1 is unicuspidate.
Kristofer M. Helgen, C. Miguel Pinto, Roland Kays, Lauren E. Helgen, Mirian T. N. Tsuchiya, Aleta Quinn, Don E. Wilson, Jesús E. Maldonado
Zookeys
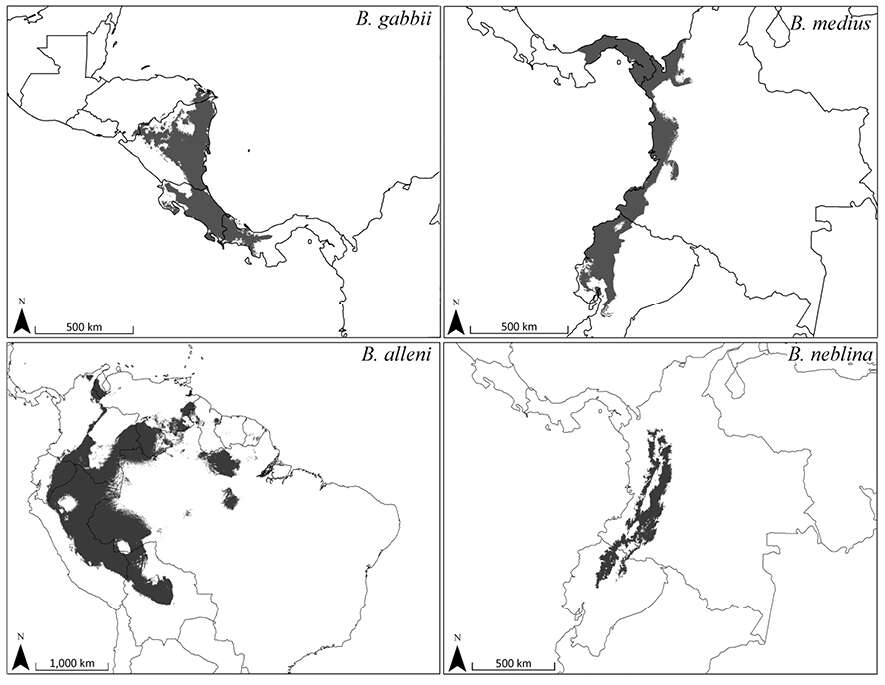
Figure 12.Predicted distribution for Bassaricyon species based on bioclimatic models. To create these binary maps we used the average minimum training presence for 10 test models as our cutoff. In addition, we excluded areas of high probability that were outside of the known range of the species if they were separated by unsuitable habitat.
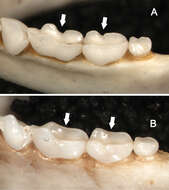
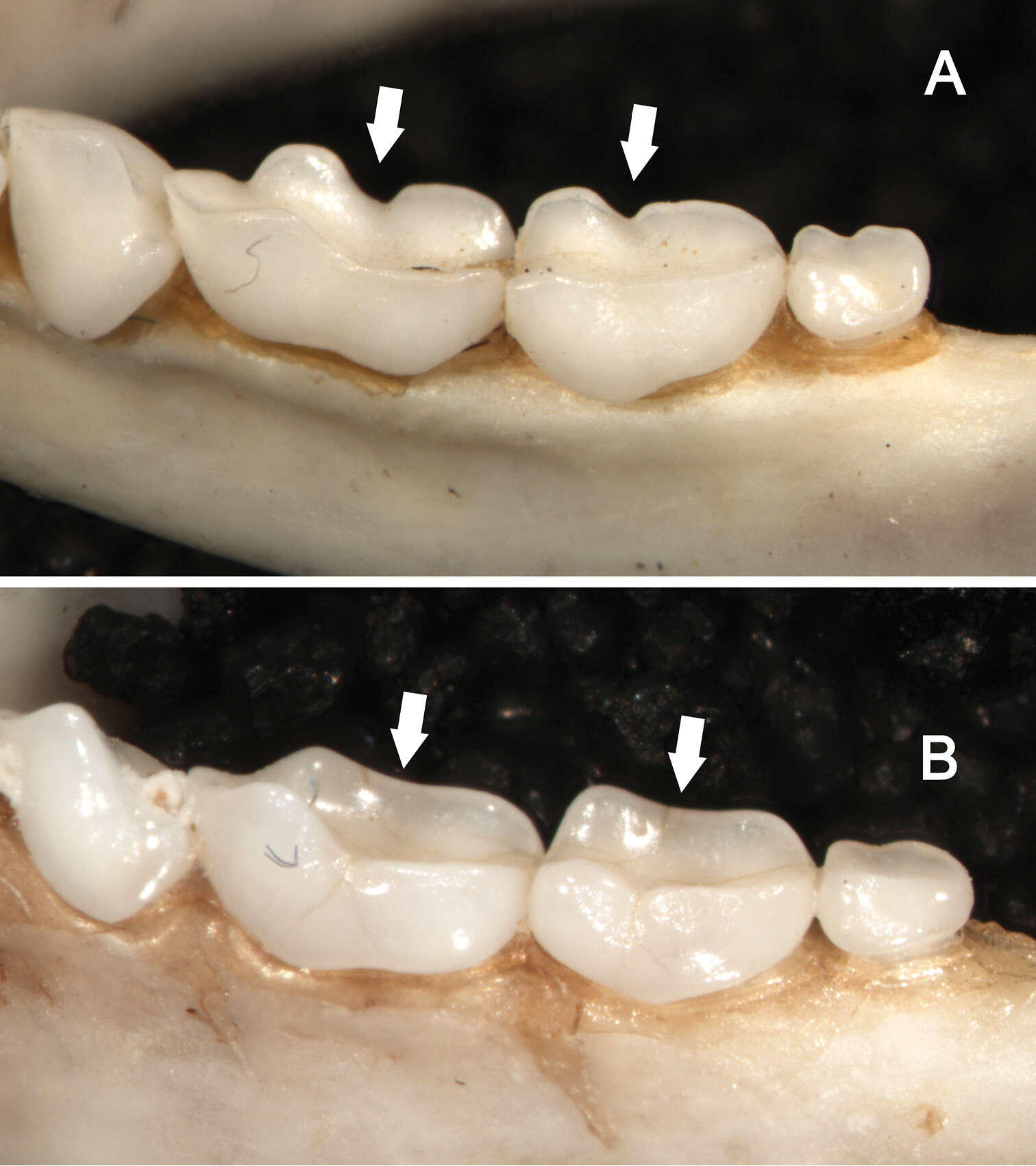
Figure 9.Dorsolateral views of the left mandibular toothrows in Sturnira bakeri (A, QCAZ 14635 ♀) and Sturnira burtonlimi (B, ROM 104294 ♂), illustrating taxonomic differences in the shape of the metaconid and entoconid of m1 and m2. In Sturnira bakeri, the metaconid and entoconid of m1 and m2 are well defined and separated by a deep notch (arrows). In Sturnira burtonlimi, however, the metaconid and entoconid of m1 and m2 are poorly defined and are not separated by a deep notch (arrows).