“Genus MACROPYXIS new genus
Derivation of name — Greek macro-, large; Greek and Latin Pyxis, a small box or casket (feminine gender).
Type species — Macrocypris sapeloensis DARBY, 1965.
Species included.—Listed by original binomen:
Macrocypris sapeloensis Darby, 1965; Holocene, northwestern Atlantic Ocean.
Macropyxis adrecta n. sp.; Holocene, equatorial Atlantic Ocean.
Macrocypris adriatica Breman, 1975; Pleistocene and Holocene, Adriatic Sea and Italy.
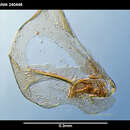
Macropyxis adunca n. sp.; Holocene, South Atlantic Ocean.
Macropyxis amanda n. sp.; Holocene, equatorial Atlantic Ocean.
Macropyxis amoena n. sp.; Holocene, Strait of Magellan and southeastern Pacific.
Macropyxis antonbruunae n. sp.; Holocene, southeastern Pacific Ocean.
Macropyxis arta n. sp.; Holocene, equatorial Atlantic Ocean.
Macropyxis audens n. sp.; Holocene, equatorial Atlantic Ocean.
Macrocypris bathyalensis Hulings, 1967a; Holocene, North Atlantic Ocean.
Macropyxis eltaninaen. sp.; Holocene, southeastern Pacific Ocean.
Macropyxis improceran. sp.; Holocene, Gulf of Mexico.
Macropyxis kaesleri n. sp.; Holocene, equatorial eastern Pacific.
Macropyxis kalbin. sp.; Holocene, equatorial eastern Pacific.
Macropyxis kornickeri n. sp.; Holocene, equatorial West Atlantic.
Macropyxis labutisi n. sp.; Holocene, northwestern Atlantic.
Macrocypris longana van den Bold, 1960; Eocene, Trinidad.
Macrocypris rhodanavan den Bold, 1960; Eocene–Miocene, Trinidad and Cuba.
Macrocypris similis Brady, 1880; Holocene, equatorial western Atlantic Ocean.
Macropyxis simulans n. sp.; Holocene, Gulf of Mexico.
Macropyxis steineckin. sp.; Miocene, Jamaica.
Macrocypris tenuicauda Brady, 1880; Holocene, northwestern Atlantic.
Macropyxis sp. 8; Holocene, southwestern Indian Ocean.
Macropyxis sp. 9; Holocene, Gulf of Gascony.
Macropyxis sp. 10; Miocene, Aquitaine Basin, France.
Macropyxis sp. 11; Holocene, equatorial western Atlantic Ocean.
Macropyxis sp. 12; Holocene, Gulf of Mexico.
Macropyxis sp. 13; Holocene, Mozambique Channel.
Macropyxis sp. 14; Holocene, Tasman Sea.
Macropyxis sp. 15; Holocene, Tasman Sea.
Macropyxis sp. 16; Holocene, southwestern Pacific Ocean.
Macropyxis sp. 17; Holocene, equatorial East Pacific Ocean.
Macropyxis sp. 18; Holocene, equatorial Atlantic Ocean.
Macropyxis sp. 20; Holocene, southeastern Atlantic Ocean.
Macropyxis sp. 21; Holocene, southwestern Atlantic Ocean.
Macropyxis sp. 46; Holocene, southeastern Pacific Ocean.
Geologic range — Eocene to Holocene.
Geographic range — Cosmopolitan, known composite live depth range 49 to 6134 m, dead specimens as shallow as 9.3 m.
Diagnosis — Carapace large, thick-shelled, smooth, translucent to opaque white without patch pattern. Elongate-ovate in lateral view; anterior margin broadly and evenly rounded; dorsal margin very broadly and gently arched, without dorsal or posterodorsal angles; ventral margin straight to slightly convex, which is usually more or less sinuous, especially posteriorly, with prolonged and gently tapering posterior region ending in more or less acutely pointed to truncate posterior angle. Moderately inflated in dorsal view; margins not compressed; anterior and posterior ends bluntly rounded; lateral margins gently curving to nearly parallel; valves distinctly asymmetrical. Hinge thick and sturdily developed; zone of concrescence moderately wide to very wide, even; radial pore canals more or less regularly and complexly branching; vestibule not constricted; few, small sensillum pores with short, smooth, simple, unbranched setae; adductor muscle-scar group fairly small in proportion to carapace size, asymmetrical in the two valves, usually diagonally oriented.
Antennules of moderately robust proportions, tapering, with most podomeres longer than wide, with short, thick setae; podomeres II and III more or less flexibly articulated. Antenna robust with numerous, fairly long, thick terminal claws; very long sensory aesthetascs on podomere II; four or more long swimming setae on podomere III; anterodorsal setae of podomere IV very long. Podomeres I and II of maxillule palp completely fused without suture; the anterodistal setae of these two podomeres remain distinct.
Palp of male right fifth limb forms a fairly large, thick, right-angled to recurved hook; left limb more tightly recurved; two small to large, fairly slender, tapering ventral pegs and one small ventral seta; proximally sensory seta and very conspicuous terminal sensory disk present, but no dorsal seta. Palp of female fifth limb slender to robust, with three terminal setae of graduated lengths, the dorsal on the shortest and the ventral one usually the longest. Sixth leg with two distal claws of equal or nearly equal size. Seventh limb with well-feathered, short to moderately long, reflexed seta and may be weakly pinnate. Furca small to fairly large, consisting of two long, cylindrical proximal rods, each bearing a cluster of up to four very tiny to medium-sized proximal setae located near the base, in the middle, or near the end of the rod and each terminating with a long, flexible, tapering seta; often dimorphic, usually somewhat larger in males but otherwise of similar construction to females, but occasionally absent in males, in which case it is represented only by a vestigial nub bearing four tiny proximal setae. Male hemipenis roughly subtriangular to rhomboidal to ovate in outline, with straight dorsal margin and acute to obtuse anterior angle; copulatory rod short, nearly straight to weakly curved. Zenker;s organ with medium-sized to long muscularized portion and a small to large spherical terminal bulb; vas deferens somewhat thickened, coiled in an irregular disk. A tight, regular spiral, or a tangled knot of 4 to 10 or more coils. The strands of sperm may also be coiled in a long tight spiral of 10 or more coils like a telephone cord before entering the terminal bulb.
Affinities – Macropyxis includes many species formerly assigned to Macrocypris, and most general statements about that genus by most previous authors actually apply to Macropyxis. Certain undescribed Australian species referred to Macrocyprissa (McKenzie, 1967a, 1974, 1976; McKenzie and Peypouquet, 1984) probably belong to Macropyxis (or possibly Macromckenziea), and the generalizations of those authors about “Macrocypris” should be regarded as referring to Macropyxis.
Macropyxis is represented by numerous species in bathyal and abyssal environments of all oceans and is far more diverse than Macrocypris or Macromckenziea. It is, in fact, the typical representative of the Macrocyprididae in psychrospheric faunas.
Remarks – Although the branching radial pore canals are the most visible, characteristic attribute of this genus, they show considerable interspecific and intraspecific variability as well. A few specimens in any species population have narrower fused marginal zones with correspondingly shorter, less elaborately branching radial pore canals than usueal for the species. Perhaps some of these specimens had molted only recently and had not completed calcification of the carapace at the time of collection. Of course this phenomenon is not, by any means, restricted to Macropyxis; a few thin-edged, juvenile-appearing adults may be found in large populations of any other genus and species of Macrocyprididae. It deserves special mention for Macropyxis only because failure to recognize this variability in marginal development may hinder identification of the genus as well as the species. Generally, the most completely developed specimens, with the most complexly branching radial pore canals, should be selected for comparison and identification.
Many species of Macropyxis also show moderate variability in the degree of prolongation of the posterior carapace angle, with accompanying variability of the exact shape of the posterior and posteroventral margin. In large populations at one station this intraspecific variation is readily assessed and recognized. For small populations or for single specimens from isolated station, though, the evaluation of shape may be equivocal, especially as the shape of the posterior angel also varies with sex and sometimes with depth or geographic distance. Thus, isolated specimens of some species may be difficult to identify reliably from carapace shape alone. Several such specimens are here treated conservatively as separate species in open nomenclature.” (Maddocks 1990, p.57-58)