
Eukariote is lede van die domein Eukaryota: organismes wat organelle binne-in hul selle bevat, soos die selkern, golgi-apparaat, mitochondrion en in plante, chloroplaste. Die domein bevat alle plante, diere, alge, swamme en protiste.
In taksonomie word eukariote beskou as een van die drie biologiese domeine waarin lewe verdeel kan word. Die ander twee domeine, Archaea en Bacteria, word saam "prokariote" genoem.
Die verwantskap tussen die eukariote is steeds 'n onderwerp van navorsing. Hierdie is 'n vereenvoudigde kladogram wat die belangrikste groepe toon:
LeweGlaucophyta
Rhodophyta
Opisthokonta (diere, swamme)
Eukariote is lede van die domein Eukaryota: organismes wat organelle binne-in hul selle bevat, soos die selkern, golgi-apparaat, mitochondrion en in plante, chloroplaste. Die domein bevat alle plante, diere, alge, swamme en protiste.
In taksonomie word eukariote beskou as een van die drie biologiese domeine waarin lewe verdeel kan word. Die ander twee domeine, Archaea en Bacteria, word saam "prokariote" genoem.
Les célules eucariotes son les que tienen el so material hereditariu fundamental (la información xenética) zarráu nuna doble membrana, la envolvedura nuclear, que rodea'l nucleu celular.
Les célules eucariotes caracterícense por presentar fenómenos de mitosis y meiosis, procesos de división celular que tienen comu finalidá la formación de nuevos individuos y la continuación de la especie.
Dientru del grupu eucariota, faese diferencia entre célules eucariotes animales y célules eucariotes vexetales.
Les eucariotes animales, son les célules que presenten tolos animales, nun puen vivir aislaes y xúntense pa formar texíos. Asina tenemos los diferentes texíos nos animales, texíu nerviosu, texíu óseo, texíu muscular... Caracterícense tamién pola presencia d'orgánulos ya coñecíos comu puen ser retículu endoplasmáticu,peroxisomes, ribosomes, nucleolu y mitocondries. Les célules eucariotes animales enxamás tendrán cloroplastos, dexando estos en posesión delles célules eucariotes vexetales.
Les célules eucariotes animales, presenten fenómenos de divisíon o reproducción asexual, tamién coñecía col nome de Mitosis. Esta división ye una division vexetativa, nos organismos pluricelulares, tien la finalidá de reparar texíos dañaos, o l'aumentu dellos mesmos.
La Meiosis, ye'l procesu de reproducción sexual que ocurre solamente nes célules xerminales, tamién coñecías col nome de gametos. Tres un proceso de diferenciación, y tres varies divisiones, fórmense los gametos, que xuntándose los femeninos y los masculinos, lleven a la formación d'una célula diploide o cigotu.
Les célules vexetales, por otra parte, presenten les mesmes estructures que les célules animales. Nesti casu, la célula vexetal, poseerá cloroplastos, orgánulos fotosintetizadores con pigmentu clorofila que intervendrán nos procesos anabólicos de la célula. Les célules vextales caracterícense tamién por presentar órganulos mui voluminosos, nomaos Vacuoles. Nes vacuoles,almacénense toles sustancies de provechu pala célula, como por exiemplu l'almidón, o los polisacáridos.

Eukariotlar və ya Nüvəlilər (lat. Eucaryota yun. εύ- — yaxşı və κάρυον — nüvə) — Hüceyrələrində nüvə olan canlı orqanizmlərin domeni (aləmüstü). Bakteriyalar və Arxeyalardan başqa bütün orqanizmlər nüvəlidir.
İşıq-optiki və elektron mikroskopları vasitəsilə müxtəlif bir və çoxhüceyrəli orqanizmlərin tədqiqi göstərmişdir ki, hüceyrələr öz quruluşlarına görə iki qrupa bölünürlər.
Bir qrup – bakteriyalar və goy-yaşıl yosunlar təşkil edirlər. Bu orqanizmlər nisbətən sadə hüceyrə quruluşuna malik olurlar. Bunları prokariot (nüvəsiz) adlandırırlar, çünki onlarda formalaşmış nüvə (yunanca "karyon"-nüvə) və həm də orqanella və ya orqanoid adlanan bir çox quruluşlar yoxdur. Digər qrupa bütün yerdə qalan orqanizmlər, birhüceyrəli göy-yaşıl yosunlardan və ibtidailərdən başlamış ali çiçəkli bitkilər, məməlilər, o cümlədən insan aiddir. Onların hüceyrələri mürəkkəb olub nüvəli eurokariot adlanır.Bu hüceyrələr nüvəyə və spesifik funksiyaları yerinə yetirən orqanellalara malik olurlar.
Prokariot hüceyrələr tək hüceyrə membranına malik olan xırda və sadə quruluşlu hüceyrələrdir. Onların orta ölçüsü 5 mkm olur. Onlarda, adətən, membran quruluşuna malik orqanellalar yoxdur, məs., mitoxondirilər və endoplazmatik tordakı kimi. Hüceyrə nüvəsinin əvəzinə onun ekvivalenti (nukleoid) olur ki, bu da sıx spiral şəkilində burulmuş yeganə bir DNT molekulundan ibarətdir. Bundan başqa bakteriyalar eukariotların nüvə xarici DNT- lərinə oxşar xırda plazmid formalı DNT-lərdən ibarət ola bilir. Fotosintez qabiliyyətinə malik olan prokariot hüceyrələrdə (göy-yaşıl yosunlar, yaşıl və al-qırmızı bakteriyalar) membranların müxtəlif iri qabarıqları tilakoidlər vardır. Bunlar öz funksiyalarına görə eurokariotların plastidlərinə uyğundurlar. Bu tilakoidlər funksional cəhətdən mitoxondiriləri əvəz edirlər. Prokariotlar yer üzündə maddə enerjinin bioloji çevrilmələrində mühüm rol oynayırlar. Fotosintezedici bakteriyalar günəş enerjisini udaraq onu karbohidratların və hüceyrələrin digər komponentlərinin sintezi üçün istifadə edilir ki, bunlar da öz növbəsində digər orqanizmlər üçün qida kimi istifadə edilir. Bəzi bakteriyalar atmosferdən molekulyar azotu fiksə edərək bioloji faydalı azottərkibli birləşmələr əmələ gətirirlər.
Çox güman ki, prokariotlar bioloji tekamül prosesində eukariotlardan əvvəl əmələ gəlmişlər.
Eukariot hüceyrələr prokariotlara nisbətən daha iri və mürəkkəbdirlər. Eukariot hüceyrələrin orta ölçüsü 13 mkm-ə yaxındır.Onların həcmləri prokariot hüceyrələrin həcmindən 1000-10000 dəfə böyük olur. Hüceyrə daxili membranlarla müxtəlif kompartmentlərə ayrılmışdır. Orqanoidlərin üç növü: hüceyrə nüvəsi, mitoxondriya və plastidlər protoplazmadan iki membrandan ibarət qişa ilə ayrılmışdır.
Eukariotlar və ya Nüvəlilər (lat. Eucaryota yun. εύ- — yaxşı və κάρυον — nüvə) — Hüceyrələrində nüvə olan canlı orqanizmlərin domeni (aləmüstü). Bakteriyalar və Arxeyalardan başqa bütün orqanizmlər nüvəlidir.
İşıq-optiki və elektron mikroskopları vasitəsilə müxtəlif bir və çoxhüceyrəli orqanizmlərin tədqiqi göstərmişdir ki, hüceyrələr öz quruluşlarına görə iki qrupa bölünürlər.
Bir qrup – bakteriyalar və goy-yaşıl yosunlar təşkil edirlər. Bu orqanizmlər nisbətən sadə hüceyrə quruluşuna malik olurlar. Bunları prokariot (nüvəsiz) adlandırırlar, çünki onlarda formalaşmış nüvə (yunanca "karyon"-nüvə) və həm də orqanella və ya orqanoid adlanan bir çox quruluşlar yoxdur. Digər qrupa bütün yerdə qalan orqanizmlər, birhüceyrəli göy-yaşıl yosunlardan və ibtidailərdən başlamış ali çiçəkli bitkilər, məməlilər, o cümlədən insan aiddir. Onların hüceyrələri mürəkkəb olub nüvəli eurokariot adlanır.Bu hüceyrələr nüvəyə və spesifik funksiyaları yerinə yetirən orqanellalara malik olurlar.
Prokariot hüceyrələr tək hüceyrə membranına malik olan xırda və sadə quruluşlu hüceyrələrdir. Onların orta ölçüsü 5 mkm olur. Onlarda, adətən, membran quruluşuna malik orqanellalar yoxdur, məs., mitoxondirilər və endoplazmatik tordakı kimi. Hüceyrə nüvəsinin əvəzinə onun ekvivalenti (nukleoid) olur ki, bu da sıx spiral şəkilində burulmuş yeganə bir DNT molekulundan ibarətdir. Bundan başqa bakteriyalar eukariotların nüvə xarici DNT- lərinə oxşar xırda plazmid formalı DNT-lərdən ibarət ola bilir. Fotosintez qabiliyyətinə malik olan prokariot hüceyrələrdə (göy-yaşıl yosunlar, yaşıl və al-qırmızı bakteriyalar) membranların müxtəlif iri qabarıqları tilakoidlər vardır. Bunlar öz funksiyalarına görə eurokariotların plastidlərinə uyğundurlar. Bu tilakoidlər funksional cəhətdən mitoxondiriləri əvəz edirlər. Prokariotlar yer üzündə maddə enerjinin bioloji çevrilmələrində mühüm rol oynayırlar. Fotosintezedici bakteriyalar günəş enerjisini udaraq onu karbohidratların və hüceyrələrin digər komponentlərinin sintezi üçün istifadə edilir ki, bunlar da öz növbəsində digər orqanizmlər üçün qida kimi istifadə edilir. Bəzi bakteriyalar atmosferdən molekulyar azotu fiksə edərək bioloji faydalı azottərkibli birləşmələr əmələ gətirirlər.
Çox güman ki, prokariotlar bioloji tekamül prosesində eukariotlardan əvvəl əmələ gəlmişlər.
Eukariot hüceyrələr prokariotlara nisbətən daha iri və mürəkkəbdirlər. Eukariot hüceyrələrin orta ölçüsü 13 mkm-ə yaxındır.Onların həcmləri prokariot hüceyrələrin həcmindən 1000-10000 dəfə böyük olur. Hüceyrə daxili membranlarla müxtəlif kompartmentlərə ayrılmışdır. Orqanoidlərin üç növü: hüceyrə nüvəsi, mitoxondriya və plastidlər protoplazmadan iki membrandan ibarət qişa ilə ayrılmışdır.
An eukarioted (eus ar gregaj eu, gwir ha karuon, kraoñenn, maen frouezh[1]) zo anezho ur strollad bevien unkellig pe lieskellig a dermener diouzh framm o c'helligoù. Unan eus tri damani ar vuhez eo an eukarioted (gant an archaebacteria hag an eubacteria).
An eukarioted o deus, er c'hontrol d'ar prokarioted (archaebacteria hag eubacteria) :
A-raok e veze rannet an eukarioted e 4 riezad : Animalia, Fungi, Plantae ha Protista (unkelligek anezho). Ma oa sklaer termenadur an teir riezad kentañ, ne oa ket ar memes tra evit riezad ar protisted a yae d'he ober « kement ha na oa ket » loened, plant pe foue.
E-kerzh ar bloavezhioù diwezhañ, an araokadennoù e « reizhadoniezh » o deus roet an tu, daoust ma n'eo ket anavet an holl, da vrastresañ ur wezenn filogenetek eus an eukarioted. Unan eus an dezastum diwezhañ e galleg eo an hini kinniget gant Guillaume Lecointre ha Herve ar Gwiader e Rummatadur filogenetek ar bevien embannet gant Belin.
An eukarioted (eus ar gregaj eu, gwir ha karuon, kraoñenn, maen frouezh) zo anezho ur strollad bevien unkellig pe lieskellig a dermener diouzh framm o c'helligoù. Unan eus tri damani ar vuhez eo an eukarioted (gant an archaebacteria hag an eubacteria).
An eukarioted o deus, er c'hontrol d'ar prokarioted (archaebacteria hag eubacteria) :
organitoù hag a rann diabarzh ar gellig e takadoù, dezho kefridioù resis, evel nukleus bevennet (ennañ an TDN), ar mitokondriennoù, pe ar reticulum endoplasmek, Ardivink Golgi, ar lisosomoù, ar peroksisomoù, ar c'hloroplastoù hag ar vacuolennoù evit a sell ar plant ; Un TDN fetisaet e stumm kromozomoù e-pad ar rannadur kelligel ; ur seksualegezh wirion, ma vez degaset ul lodenn gevatal eus an dafar genetek gant hiniennoù an div jener.
A-raok e veze rannet an eukarioted e 4 riezad : Animalia, Fungi, Plantae ha Protista (unkelligek anezho). Ma oa sklaer termenadur an teir riezad kentañ, ne oa ket ar memes tra evit riezad ar protisted a yae d'he ober « kement ha na oa ket » loened, plant pe foue.
E-kerzh ar bloavezhioù diwezhañ, an araokadennoù e « reizhadoniezh » o deus roet an tu, daoust ma n'eo ket anavet an holl, da vrastresañ ur wezenn filogenetek eus an eukarioted. Unan eus an dezastum diwezhañ e galleg eo an hini kinniget gant Guillaume Lecointre ha Herve ar Gwiader e Rummatadur filogenetek ar bevien embannet gant Belin.
Els eucariotes (de noms científics Eucaryotae, Eukaryotae, Eukarya o Eucarya) és, en taxonomia i histologia, el domini d'organismes cel·lulars amb nucli diferenciat,[1] i altres orgànuls tancats dins membranes biològiques, contraposats així als procariotes. Els eucariotes pertanyen al tàxon Eukarya o Eukaryota. L'estructura que defineix l'estructura de la membrana que fa que les cèl·lules eucariotes estiguin a part de les cèl·lules procariòtiques (bacteris i Archaea) és el nucli cel·lular, el qual conté el material genètic, tancat dins l'embolcall nuclear.[2][3][4] La presència d'un nucli dóna als eucariotes el seu nom, el qual prové del grec: ευ (eu, 'ben') i κάρυον (karyon, 'nou' o 'pinyol').[5] Les cèl·lules eucariotes també contenen altres orgànuls amb membrana com els mitocondris o els aparells de Golgi. A més, les plantes i les algues contenen cloroplasts. Molts organismes unicel·lulars són eucariotes, com ho són els protozoa. Tots els organismes multicel·lulars són eucariotes, incloent-hi animals, plantes i fongs.
Als organismes formats per cèl·lules eucariotes se'ls denomina eucarionts.
Les anàlisis seqüenciadores dels ribosomes avalen la teoria que aquestes seqüències dels eucariotes, les principals en la taxonomia genètica, són més properes als arqueobacteris que no pas als bacteris i que van compartir un avantpassat comú amb els primers durant més temps que els segons. Tanmateix, altres grups de gens estan més relacionats amb els eubacteris i encara d'altres semblen ser una diversificació anterior als dos altres dominis. Això es deuria a l'elevada transferència horitzontal de gens original que es va donar en el brou primitiu.
La divisió cel·lular en els eucariotes difereix de la des procariotes. Hi ha dos tipus de processos de divisió. En la mitosi, una cèl·lula es divideix per a produir dues cèl·lules genèticament idèntiques. En la meiosi, la qual es requereix en la reproducció sexual, una cèl·lula diploide experimenta recombinació: de cada parell de cromosomes parentals, en resulten quatre cèl·lules haploides (gamets).
El domini Eukaryota sembla ser monofilètic. Els dos altres dominis, Eubacteria i Archaea, són procariotes. Els eucariotes representen una petita minoria de tots els éssers vius;[6] fins i tot, en el cos humà hi ha 10 cops més microbis que cèl·lules humanes.[7]
També es denominen eucariotes les cèl·lules que tenen el seu material hereditari fonamental (la seva informació genètica, ADN) envoltat per d'una membrana cel·lular doble, l'embolcall nuclear, que delimita un nucli cel·lular.[8]
L'alternativa a l'organització eucariòtica de la cèl·lula l'oferix l'anomenada cèl·lula procariota. En aquestes cèl·lules, el material hereditari apareix més o menys dispers en el citoplasma. Les cèl·lules eucariotes no tenen un compartiment al voltant de la membrana plàsmica, com el que tenen les cèl·lules procariotes.
A diferència de les cèl·lules procariotes, les eucariotes presenten un citoplasma molt compartimentat, amb orgànuls separats o interconnectats, limitats per membranes biològiques que són de la mateixa naturalesa essencial que la membrana plasmàtica. El nucli és només el més notable i característic dels compartiments en què es divideix el protoplasma, és a dir, la part activa de la cèl·lula. En el protoplasma, distingim tres components principals, a saber: la membrana plasmàtica, el nucli i el citoplasma, constituït per tota la resta. Les cèl·lules eucariotes estan dotades d'un citoesquelet complex, molt estructurat i dinàmic, format per microtúbuls i diversos filaments proteics. A més, pot haver-hi paret cel·lular, que és el típic de plantes, fongs i protoctists pluricel·lulars, o algun altre tipus de recobriment extern a la membrana.
Les cèl·lules eucariotes contenen en principi mitocondris, orgànuls derivats per endosimbiosi de certs eubacteris, la qual cosa els dota de la capacitat de desenvolupar un metabolisme aerobi. Tanmateix, en alguns eucariotes del regne protoctists, els mitocondris han desaparegut secundàriament en el curs de l'evolució, en general derivant a altres orgànuls, com els hidrogenosomes.
Alguns eucariotes realitzen la fotosíntesi, gràcies a la presència en el seu citoplasma d'orgànuls anomenats cloroplasts, els quals deriven per endosimbiosi d'eubacteris del grup denominat cianobacteris (algues blaves).
Encara que demostren una diversitat increïble en la seua forma, comparteixen les característiques fonamentals de la seua organització cel·lular, a dalt resumides, i una gran homogeneïtat quant a la seua bioquímica (composició) i metabolisme, que contrasta amb la immensa heterogeneïtat que en aquest terreny presenten els procarionts (bacteris sensu lato).
Els organismes eucariotes formen el domini Eukarya, que inclou els organismes més coneguts, repartits en quatre regnes: Animalia (animals), Plantae (plantes), Fungi (fongs) i Protista. Inclouen la gran majoria dels organismes extints morfològicament recognoscibles que estudien els paleontòlegs. Els exemples de la disparitat eucariòtica van des d'un dinoflagel·lat (un protist unicel·lular fotosintetitzador), un arbre com la sequoia, un calamar, o un xanglot de bolets (òrgans reproductius de fongs), cadascun amb cèl·lules distintes i, en el cas dels pluricel·lulars, sovint molt variades.
Els eucariotes (de noms científics Eucaryotae, Eukaryotae, Eukarya o Eucarya) és, en taxonomia i histologia, el domini d'organismes cel·lulars amb nucli diferenciat, i altres orgànuls tancats dins membranes biològiques, contraposats així als procariotes. Els eucariotes pertanyen al tàxon Eukarya o Eukaryota. L'estructura que defineix l'estructura de la membrana que fa que les cèl·lules eucariotes estiguin a part de les cèl·lules procariòtiques (bacteris i Archaea) és el nucli cel·lular, el qual conté el material genètic, tancat dins l'embolcall nuclear. La presència d'un nucli dóna als eucariotes el seu nom, el qual prové del grec: ευ (eu, 'ben') i κάρυον (karyon, 'nou' o 'pinyol'). Les cèl·lules eucariotes també contenen altres orgànuls amb membrana com els mitocondris o els aparells de Golgi. A més, les plantes i les algues contenen cloroplasts. Molts organismes unicel·lulars són eucariotes, com ho són els protozoa. Tots els organismes multicel·lulars són eucariotes, incloent-hi animals, plantes i fongs.
Als organismes formats per cèl·lules eucariotes se'ls denomina eucarionts.
Les anàlisis seqüenciadores dels ribosomes avalen la teoria que aquestes seqüències dels eucariotes, les principals en la taxonomia genètica, són més properes als arqueobacteris que no pas als bacteris i que van compartir un avantpassat comú amb els primers durant més temps que els segons. Tanmateix, altres grups de gens estan més relacionats amb els eubacteris i encara d'altres semblen ser una diversificació anterior als dos altres dominis. Això es deuria a l'elevada transferència horitzontal de gens original que es va donar en el brou primitiu.
La divisió cel·lular en els eucariotes difereix de la des procariotes. Hi ha dos tipus de processos de divisió. En la mitosi, una cèl·lula es divideix per a produir dues cèl·lules genèticament idèntiques. En la meiosi, la qual es requereix en la reproducció sexual, una cèl·lula diploide experimenta recombinació: de cada parell de cromosomes parentals, en resulten quatre cèl·lules haploides (gamets).
El domini Eukaryota sembla ser monofilètic. Els dos altres dominis, Eubacteria i Archaea, són procariotes. Els eucariotes representen una petita minoria de tots els éssers vius; fins i tot, en el cos humà hi ha 10 cops més microbis que cèl·lules humanes.
Organeb gyda chelloedd cymhleth yw ewcaryot. Mae gan gelloedd ewcaryotau gnewyllyn sy'n cynnwys y cromosomau. Mae'r celloedd yn rhannu drwy feiosis neu fitosis.
Mae'r ewcaryotau'n ffurfio un o'r tri pharth o organebau byw; y bacteria a'r archaea yw'r lleill. Rhennir yr ewcaryotau yn bedair teyrnas yn draddodiadol: anifeiliaid, planhigion, ffyngau a phrotistiaid. Holltir y protistiaid yn sawl grŵp gwahanol gan lawer o dacsonomegwyr modern.
Rhannau celloedd ewcaryotig:
Organeb gyda chelloedd cymhleth yw ewcaryot. Mae gan gelloedd ewcaryotau gnewyllyn sy'n cynnwys y cromosomau. Mae'r celloedd yn rhannu drwy feiosis neu fitosis.
Mae'r ewcaryotau'n ffurfio un o'r tri pharth o organebau byw; y bacteria a'r archaea yw'r lleill. Rhennir yr ewcaryotau yn bedair teyrnas yn draddodiadol: anifeiliaid, planhigion, ffyngau a phrotistiaid. Holltir y protistiaid yn sawl grŵp gwahanol gan lawer o dacsonomegwyr modern.
Eukaryota (též Eukarya či česky jaderní) je velká skupina (doména) jednobuněčných a mnohobuněčných organismů, jako jsou například živočichové, rostliny, houby nebo prvoci. Eukaryotická buňka na rozdíl od prokaryotní obsahuje pravé buněčné jádro a množství dalších organel oddělených membránou od okolí. Tyto struktury rozdělují buňku na mnoho menších oddílů (kompartmentů). Eukaryontní organismy mají množství jiných unikátních znaků, jako je například specifická struktura bičíků a řasinek, existence cytoskeletu, určité odchylky ve struktuře genomu (např. DNA rozdělená do jednotlivých chromozomů, přítomnost intronů ve větším množství), schopnost pohlavního rozmnožování a mnohá další metabolická a biochemická specifika.
K eukaryontním organismům se řadí všechny buněčné organismy vyjma bakterií a archeí, tedy prokaryot. Klasifikace eukaryot se v poslední době radikálně mění. Říše rostliny, houby, živočichové a protista se v současnosti nahrazují několika přirozenými skupinami, jako jsou Amoebozoa, Opisthokonta, Rhizaria, Plantae (rostliny), Chromalveolata a Excavata. Odhady týkající se počtu druhů eukaryot se pohybují mezi 5 a 30 miliony,[1][2] popsáno bylo méně než dva miliony druhů.[2]
Vznik eukaryotických buněk (a potažmo tedy eukaryot) je významný milník v evoluční historii života, je však zahalen nejistotou. Někdy se datuje do doby před 1,8 – 1,3 miliardami lety,[3] tedy o mnoho dříve před kambrijskou a ediakarskou diverzifikací života. Jindy se dokonce uvádí doba před dvěma miliardami let[5] či více.[6] Z těchto období naší geologické historie je k dispozici jen málo fosilních nálezů eukaryot, i když výjimky existují: byly popsány starohorní fosilní nálezy protistů, které jsou označovány za časné zástupce eukaryot.[3] Také některé nálezy řas (Grypania) jsou podle jejich objevitelů až 2,1 miliardy let staré.[4] Stopy po existenci eukaryot byly dokonce nalezeny i v australských břidlicích starých 2,7 miliardy let.[7] I přes tyto nálezy se hlavní současné linie eukaryot pravděpodobně oddělily až ke konci starohor, konkrétně v období mezi 1,3 – 0,72 miliardami let. Fosílie časných eukaryot, jako je například Shuiyousphaeridium macroreticulatum, může být obtížné odlišit od prokaryotických buněk. U tohoto domnělého eukaryota se při jeho řazení například vycházelo z velkých rozměrů buňky, pevné stěny a výrůstků na jeho povrchu.[3] Většinu nálezů ze starohor však nejsou vědci schopni zařadit do konkrétnější skupiny eukaryot. Kvalitní fosílie eukaryot se začínají objevovat až v ediakaru a kambriu, tedy v období asi před půl miliardou let. V ediakaru byly nalezeny vyjma zelených řas také zvláštní planktonní organismy ze skupiny Acritarcha.[8]
Otázka vzniku eukaryotické buňky (eukaryogeneze), která je charakteristická množstvím vnitřních membrán a složitými organelami (mitochondrie), není zcela vyjasněna. Dnes se zdá, že ke vzniku eukaryot přispěly jak archeální, tak bakteriální genomy.[9] Jedním ze světlých míst je vznik mitochondrií. V této organele byla totiž nalezena DNA, která vykazuje značnou příbuznost s bakteriemi z řádu Rickettsiales. Endosymbiotická teorie tak oprávněně tvrdí, že mitochondrie vznikly právě pohlcením Rickettsií jinou buňkou a mitochondrie jsou potomci těchto bakterií.[9]
To však neznamená, že pouhý vznik mitochondrií znamenal vznik eukaryot v dnešním slova smyslu. Eukaryogenezi se snaží dále vysvětlit více než 20 hypotéz.[10] Podle archeální hypotézy vznikla protoeukaryotická buňka z buňky jisté archebakterie a mitochondrie se vyvinuly až za určitý čas, a to pohlcením bakteriálního symbionta.[9] Detailněji se okolnostmi archeální hypotézy zabývá tzv. vodíková hypotéza, podle níž zmíněná archebakterie metabolizovala vodík, který ji právě poskytovaly v rámci své respirace endosymbiotické bakterie. Tyto bakterie následně prošly evolucí a změnily se na mitochondrie.[11]
Jiné studie však navrhují o krok složitější způsob vzniku eukaryotické buňky. Takzvané chimérické (čili fúzní) hypotézy předpokládají, že amitochondriální eukaryot (tedy předek eukaryot ještě bez mitochondrií) vznikl splynutím jedné buňky archeální a jedné bakteriální. Eukaryotický genom je podle této hypotézy rovněž mixem archeálních a bakteriálních genů. Až následně vznikly mitochondrie (a plastidy), a to dalšími endosymbiotickými událostmi.[12] Podobná teorie navrhuje jiný sled událostí: archebakterie strávila bakterii jako potravu a část bakteriální DNA se včlenila do archeální DNA.[13] Zcela jiný pohled nabízí teorie „Neomura“, která považuje archea i eukaryota za potomky určité složité grampozitivní bakterie, konkrétně jisté aktinobakterie. Tato aktinobakterie prošla značným vývojem a až později endosymbioticky získala mitochondrie.[14] Nedostatkem těchto teorií je fakt, že v současnosti neznáme žádné eukaryotické organismy s primární absencí mitochondrií, ačkoliv je tyto teorie předpokládají.[9]
U eukaryot mnohobuněčnost vznikla několikrát nezávisle na sobě, zvlášť u živočichů, rostlin, hub a mnoha dalších eukaryotických taxonů.[16] Vznik (respektive vzniky) mnohobuněčnosti byl značným evolučním úspěchem eukaryot, u něhož byla například vyřešena otázka, jak se buňky navzájem dorozumí a jak si jednotlivé buněčné typy rozdělí práci.[17] U mnohobuněčných živočichů jsou unikátní nejen geny zajišťující správný embryonální vývoj (např. Hox geny), ale i geny zajišťující komunikaci mezi buňkami. U mnohobuněčných se vyskytují i nové buněčné struktury (dezmozomy a jiné buněčné spoje).[18]
První mnohobuněční živočichové vznikli v průběhu období ediakara (někdy před 570 – 550 miliony lety),[18] jak napovídají i některá molekulárně biologická data. Zřetelnější fosilní záznam se však objevuje až v kambriu. Z této doby, označované také jako kambrická exploze druhů, pochází množství fosilních dokladů současných živočišných kmenů.[19][20] U rostlin (včetně mnohých ruduch a zelených řas) se mnohobuněčnost vyvinula hned několikrát, a to zřejmě v souvislosti s tím, že se fotosyntetizující organismy nemusí tolik pohybovat za potravou, a vznik mnohobuněčnosti je pro ně tudíž schůdnější.[21] Vývoj mnohobuněčného uspořádání se u rostlin ubíral třemi základními cestami. Některé kolonie zelených řas, jaké tvoří např. váleč koulivý (Volvox globator), se zformovaly z několika původních jedinců. Jindy se v původní buňce mnohonásobně dělilo jádro, čímž došlo ke vzniku jednobuněčných, ale vícejaderných stélek (např. rod Caulerpa). Konečně třetím způsobem vznikly mnohobuněčné rostliny tak, že se původní buňka mitoticky dělila a jednotlivé dceřiné buňky se specializovaly na určitou funkci.[22] U hub nelze pozorovat nějaký obecný trend k mnohobuněčnosti a není vlastně jasné, kolikrát v rámci hub mnohobuněčnost vznikla.[18]
Skupin, u nichž byla pozorována mnohobuněčnost, je však mnohem více. Příkladem jsou akrasie (Acrasidae), chaluhy (Phaeophyceae), řasovky (Oomycota), pravé hlenky (Eumycetozoa) a dokonce několik nálevníků (Ciliophora) tvořících plodničky.[16][21]
Buňky eukaryot mají v průměru desetkrát větší rozměry (lineární) než buňky prokaryotických organismů,[23] ačkoliv toto pravidlo platí jen zhruba. Nejmenší eukaryotické buňky má zelená řasa Ostreococcus tauri, a to přibližně jeden μm,[24] tzn. menší než např. buňka bakterie Escherichia coli.[25] Naopak známe mnohé obrovské eukaryotické buňky. Značných rozměrů dosahují zejména některé mnohojaderné buňky uvnitř těl velkých živočichů (jako např. některé nervové buňky), které jsou však neschopné samostatné existence. Také žloutek ve vejcích pštrose či vyhynulého ptáka Aepyornis dosahuje obrovských rozměrů, ačkoliv před oplozením představuje též jen jedinou buňku.[26] Pokud se však zaměříme na největší buňku schopnou samostatné existence, existují i mimořádně velké jednobuněčné organismy. Velmi velká (až jeden metr) je jednobuněčná zelená řasa rodu Caulerpa.[27] Mnohojaderná plazmodia prvoků, jako je Physarum polycephalum, mohou také dosahovat velikosti několika metrů – zaznamenáni byli jedinci uvnitř těl velkých živočichů s plochou 5,54 m2.[28]
Buňka eukaryot je radikálně odlišná od prokaryotické buňky nejen v otázce velikosti, ale vykazuje značné rozdíly i po stránce strukturní. Na rozdíl od prokaryot jsou komplexnější a tzv. kompartmentalizované, tedy rozdělené na jednotlivé kompartmenty („oddělení“). Obsahují například pravé jádro obklopené dvojitou membránou, jež odděluje DNA od okolních částí buňky. Dalším výrazným rysem je endomembránový systém, tedy skupina organel rovněž obalených membránou. K těmto patří zejména endoplazmatické retikulum, Golgiho aparát, lyzozomy či mitochondrie, případně funkční deriváty mitochondrií (hydrogenozomy, mitozomy). Evoluční novinkou je na rozdíl od prokaryot i velmi rozvinutý cytoskelet, jakási vnitřní kostra buňky tvořená především aktinem a tubulinem. Ten zajišťuje nejen pohyb buněk, ale i vnitřní transport a zastává též důležité funkce při dělení buněk. Neméně důležitou součástí jsou ribozomy, u eukaryot jsou tzv. 80S (toto číslo udává čas, za který proběhne sedimentace ribozomální makromolekuly při její ultracentrifugaci).
Mimo výše uvedené struktury, které jsou přítomné téměř ve všech eukaryotických buňkách, se v mnoha skupinách eukaryot vyvinuly i jiné specifické organely. Typickým příkladem jsou plastidy, semiautonomní organely, v nichž probíhá fotosyntéza a některé další pochody. Vyskytují se u rostlin a mnohých protistů. Vznikly pravděpodobně v evoluční historii až určitou dobu po vzniku samotné eukaryotické buňky a samotné eukaryogeneze se neúčastnily. Přesto jsou však dalším dokladem endosymbiotické teorie.[29] Na rozdíl od mitochondrií se plastidy vyvinuly u několika, často nepříbuzných skupin eukaryot. Tzv. primární plastidy však pochází ze sinic a vznikly pravděpodobně pouze jednou, a to u rostlin, které v širším pojetí zahrnují nejen zelené rostliny (Viridiplantae), ale i ruduchy (Rhodophyta) a glaukofyty (Glaucophyta).[30] Plastidy u jiných druhů fotosyntetizujících eukaryot vznikly především sekundární endosymbiózou, tedy pohlcením jedné ze skupin rostlin. Pohlcením ruduchy vznikly plastidy (někdy kvůli svému původu zvané rhodoplasty) např. u různých heterokont (Heterokonta) a rozsivek (Dinoflagellata), pohlcením zelené řasy vznikly plastidy u Chlorarachniophyta, některých krásnooček (Euglenoidea) a jedné obrněnky.[29]
Zvláště u různých jednobuněčných protistů je známo mnoho dalších unikátních organel: vystřelovací či vymršťovací útvary (tzv. extruzomy), přichycovací organely (různé stonky a přísavky), stažitelné, potravní a jiné vakuoly, a podobně. K pohybu zase slouží různé bičíky a brvy (viz kapitola pohyb). Zcela samostatnou kapitolou je anatomie mnohobuněčných organismů (viz anatomie rostlin, anatomie živočichů, anatomie hub).
Historické snahy o klasifikaci živých organismů jsou z velké části právě klasifikací eukaryot. Na nejvyšší úrovni v průběhu času docházelo k velkým změnám. Zatímco Linné rozeznával jen dvě základní říše, rostliny a živočichové, postupně se ukázalo, že jsou si tyto skupiny (a dále někteří jednobuněční a houby) v mnoha rysech vzájemně podobné a zároveň diametrálně odlišné od bakterií. Pravděpodobně prvním, kdo si toto uvědomil, byl francouzský biolog Edouard Chatton. Ten v roce 1937 navrhl dělit všechny živé organismy na dvě říše, eukaryota a prokaryota. Jeho návrh však poněkud zapadl a znovu se k němu v roce 1962 vrátili Roger Stanier a Cornelius Van Niel. Ačkoliv je v současnosti taxon prokaryota považován za zastaralý a nerespektující fylogenetický vývoj, taxon eukaryota se běžně používá.[37] Podpořili ho v roce 1990 Carl Woese, Otto Kandler a Mark Wheelis, kteří rozčlenili veškerý život do tří domén, Archaea, Bacteria a Eucarya.[38]
Tradičně se eukaryota dělila na několik říší, například podle Whittakera[39] na říše živočichové (Animalia), rostliny (Plantae), houby (Fungi) a protisté (Protista). Cavalier-Smith dále rozdělil říši protistů na dvě dílčí, a to protozoa a chromista.[40] Dnes se však prosazují zcela jinak postavené systémy.[41]
Většina dnešních systémů uznává třídoménový systém z roku 1990 a s ním spojenou existenci domény eukaryota.[42][43] Eukaryota a archea jsou společně někdy řazena do skupiny Neomura, jelikož se zdá, že jsou tyto dvě domény vzájemně příbuzné.[44][45]
Klasifikace této domény na nižších úrovních však není zcela ustálena. Zatímco se některé systémy stále drží taxonů prvoci (Protozoa), protisté (Protista) či chromisté (Chromista),[42] jiné systémy s těmito termíny vůbec či téměř neoperují. Do centra pozornosti se staví několik skupin, označovaných jako říše,[46] infraříše[47] „supergroups“[48] či „megagroups“.[5]
Velká část klasifikačních systémů nadále spoléhá, v souladu s názvoslovnými normami, na tradiční taxonomické úrovně, přičemž se je snaží naplnit co nejpřirozeněji s ohledem na skutečnou příbuznost. Omezené množství klasifikačních úrovní však neumožňuje vždy vyjádřit podřízenost, a tak přetrvávají i taxony, kladoucí na stejnou úroveň více podřízených taxonů, než pouhé sesterské skupiny a často obsahují parafyletické, případně i polyfyletické taxony. Naopak se v těchto systémech zpravidla daří na některou úroveň umístit i skupiny nevyjasněného postavení, tzv. incertae sedis a skupiny vyhynulé. Využívají zpravidla tradičně zavedené říše, třebaže si nové poznatky vynutily jejich částečné přeskupení a pozměněné naplnění.
Systémy založené na tomto tradičním přístupu jsou vhodné pro katalogizaci organismů a proto se používají v mnohých internetových katalozích, jako Catalogue of Life, Species 2000, AlgaeBase, WoRMS apod. Z pedagogických důvodů jsou vhodné pro podání základního přehledu o eukaryotních organismech, naopak nevhodné pro problematiku evoluce a vzájemné příbuznosti.
Příklad ukazuje následující tabulka (podbarvení říší odpovídá podbarvení taxoboxů):
„Říše“Jiné klasifikační systémy využívají důsledněji fylogenetický přístup a jsou proto strukturovanější a lépe respektují přirozenost (monofylii) jednotlivých skupin a kladogramy získané z fylogenetických analýz – protože zpravidla nepoužívají tradiční taxony, mají možnost vyjádřit do veliké hloubky vzájemné příbuzenské vztahy. I z tohoto důvodu však oplývají velkým množstvím incertae sedis na všech úrovních a vzhledem k dosud nedostatečnému pokrytí molekulárně genetickými daty si mnohý nový poznatek vyžádá zásadnější revizi systému.
Konkrétně u eukaryot se při fylogenetickém přístupu podařilo nastolit i vyvrátit mnoho hypotéz o příbuznosti a přirozenosti vyšších skupin, což bylo zpravidla spojeno se změnou struktury systému. Lze uvést mnoho příkladů:[49]
Klasifikace důsledně založená na fylogenetickém přístupu (v nejpřísnější podobě je to kladistika) je používána ve většině odborných článků zabývajících se příbuzenskými vztahy organismů, jejich evolucí a fylogenetickou klasifikací; pro objasňování této problematiky je vhodná i z pedagogických důvodů, nehodí se však pro představení základního přehledu eukaryotních organismů.
Takto založené klasifikační systémy jsou zpravidla v textu prezentovány odrážkovou strukturou s proměnným odsazením, ale nejčastěji graficky pomocí fylogenetických stromů.
Zjednodušený fylogenetický strom eukaryot podle současných (tj. počátek r. 2019)[49] představ (záměrně není vyznačen kořen stromu, jehož poloha není dosud definitivně vyjasněna):
Diaphoretickes rostlinyCryptista (skrytěnky, pikomonády)
Haptista (haptofyty, centrohelidní slunivky)
Stramenopiles (rozsivky, zlativky, chaluhy, řasovky…)
Discoba (krásnoočka, améboflageláti…)
„CruMs“
Genetický materiál v podobě DNA je uložen v jádře (či několika jádrech, jako u nálevníků) a v semiautonomních organelách, jako jsou mitochondrie a plastidy (vzácně též jinde, např. v nukleomorfu, zbytku jádra po endosymbiontovi). Tato DNA se následně, podobně jako u ostatních domén organismů, přepisuje do RNA a ta pak slouží buď k syntéze proteinů (mRNA), nebo má určitou katalytickou funkci jako taková (rRNA, tRNA a podobně).
V jádře tvoří DNA a některé proteiny (např. histony) tzv. chromatin, jehož jednotlivé oddělené části se nazývají chromozomy. Stáčením (spiralizací) a rozplétáním (despiralizací) chromatinu se značně mění jejich velikost. Velikost jaderného genomu (počet bází) je značně rozmanitá. Nejmenší eukaryotický genom, pouze 2,9 milionu párů bází, má vnitrobuněčná parazitická mikrosporidie jménem Encephalitozoon cuniculi,[55][56] a to proto, že u ní došlo k redukci genomu v rámci jejího parazitického způsobu života. Naopak největší genom ze všech známých eukaryot (670 miliard párů bází) má Amoeba dubia.[57] Oproti prokaryotním organizmům u eukaryot většina jejich DNA nekóduje žádný gen (a tedy ani protein). U lidí tvoří nekódující DNA asi 97 % genomu a její funkce, pokud nějaké vůbec má, není v drtivé většině případů dosud známa. Označuje se též jako junk DNA, související termíny jsou také introny či repetitivní DNA.[22]
Mitochondriální DNA a plastidová DNA jsou oproti jaderné DNA relativně malé a cirkulární a považují se za pozůstatek po endosymbiotických událostech, při nichž byl eukaryontní buňkou pohlcen prokaryotní organismus. Mitochondriální DNA má mnoho společných znaků s DNA alfa proteobakterií, plastidová vykazuje příbuznost s DNA cyanobakterií (sinic).
Metabolické pochody výrazně ovlivňuje už vyšší velikost eukaryontní buňky, která značně zvyšuje poměr objemu a plochy buňky.[22] V podstatě rozeznáváme u eukaryot dva druhy výživy: obligátní fotoautotrofii u rostlin a jiných fotosyntetizujících organismů a obligátní chemoheterotrofii u živočichů, hub a jiných organismů živících se organickým materiálem.[58] Pokud umí nějaký organismus užívat oba tyto základní typy výživy (např. mnohá krásnoočka), označuje se termínem mixotrof. Najdeme však drobné odchylky od klasických způsobů získávání živin a energie. Někteří prvoci a houby například dokáží metabolizovat i bez kyslíku.[59]
V reprodukci eukaryot hraje velkou roli nepohlavní rozmnožování, ale evolučně unikátní je zřejmě schopnost rozmnožovat se pohlavně. Poměrně často dochází k pravidelnému střídání pohlavního a nepohlavního rozmnožování (metageneze). Pouze u některých protistů, jako trubénky (Choanoflagellata), eugleny (Euglenoidea), trichomonády (Trichomonadida) a většina krytének (Arcellinida), nebyla sexuální reprodukce dosud nalezena a o její existenci můžeme jen spekulovat.[28]
Nepohlavně se množí většina protistů, ale i některé mnohobuněčné organismy. U jednobuněčných organismů je rozmnožování asociováno s mitózou. Před vlastní cytokinezí musí být replikována DNA a také se musí zmnožit organely, aby každá dceřiná buňka byla kompletní. Rozlišuje se několik typů nepohlavního rozmnožování, a to prosté dělení ve dva (u protistů nejběžnější), dále pučení (např. rournatky, Suctoria) či mnohonásobné dělení buňky za vzniku spousty dceřiných buněk (různí výtrusovci, Apicomplexa).[28] U mnohobuněčných organismů se vyskytuje široké spektrum různých druhů nepohlavního rozmnožování, jako je vegetativní rozmnožování (např. fragmentace u mnohých rostlin), tvorba spor nepohlavní cestou, partenogeneze, apomixie či též pučení.
I pohlavní rozmnožování je v rámci eukaryotické domény neobyčejně rozšířené (až na výše zmíněné výjimky). Typický scénář u mnoha prvoků i mnohobuněčných organismů vypadá takto: v procesu meiózy vznikají dva typy sexuálně diferencovaných buněk, tzv. gamety, obvykle označované jako + a – (či samčí a samičí). Pokud tyto buňky vypadají velmi podobně, pak rozmnožování je izogamické, pokud jsou odlišné, pak se mluví o anizogamii. Specifickým typem je pak oogamie, při níž je velká samičí pohlavní buňka oplodněna malou pohyblivou samčí pohlavní buňkou. Někdy splývají rovnou rodičovské buňky (gamonti), příkladem je konjugace nálevníků.[28] U některých mnohobuněčných organizmů se vyvinuly specializované pohlavní orgány.
Charakter pohybu eukaryotických organismů se do značné míry odvíjí od jejich tělní stavby. Jiný typ pohybu najdeme u jednobuněčných eukaryot, jinak se samozřejmě pohybují mnohobuněční.
U jednobuněčných organismů či jednobuněčných stádií mnohobuněčných organismů (př. spermie) hlavní roli hrají zejména řasinky či bičíky, struktury o průměru asi 0,2 mikrometru. Vlastní úzké tělo (axonema) je celé povlečeno plazmatickou membránou a nachází se tedy uvnitř buňky. Na příčném průřezu je vidět unikátní uspořádání mikrotubulů (9×2+2), při němž je jedna centrální dvojice mikrotubulů obklopena kruhovitě devíti dalšími. Tato struktura je velmi stabilní a liší se jen u několika skupin. Pohyb bičíků a řasinek spotřebovává ATP a je založen na činnosti tzv. ATPáz. Známe však i mnoho jiných struktur umožňujících pohyb samostatných buněk. Améboidní (měňavkovitý) pohyb eukaryot je založen na polymeraci aktinu a vyskytuje se například u zástupců říše Amoebozoa či také mnoha dírkonošců (Foraminifera). Charakteristický druh pohybu u krásnooček (Euglenida) zahrnující vlnivé změny těla se nazývá euglenoidní pohyb. Klouzavým pohybem, který je zřejmě založen na cíleném vylučování slizu, se mohou pohybovat například hromadinky (Gregarinidea) či kokcidie (Coccidea).[28] U mnohobuněčných organismů se rozvinuly zcela jiné druhy lokomoce, povětšinou založené na kontrakci speciálních typů buněk, a zahrnující plazení, chůzi, běh, plavání a podobně.
Eukaryotické organismy žijí téměř všude, kde je dostatečný konstantní přívod energie, a to buď sluneční záření pro autotrofy, nebo dostatek živin pro výživu heterotrofů. Extrémofilní eukaryota, podobně jako mnohé bakterie a archea, žijí i na mnoha neobvyklých stanovištích s extrémními podmínkami. Omezujícím faktorem je pro eukaryotické organismy zejména teplota prostředí, která na rozdíl od bakterií a archeí (ani u většiny termofilních eukaryot) nesmí přesáhnout 70 °C.[60]
Výčet extrémních prostředí je poměrně dlouhý. Známe anaerobní eukaryota (mnozí protisté s hydrogenozomy, kvasinky), termofily (např. ruducha Cyanidium caldarium, termofilní houby z kompostů, někteří mnohoštětinatci), chladnomilná eukaryota (psychrofilní protisté z arktických a antarktických moří), acidofilové (Cyanidium caldarium a několik hub žijících v pH 0), alkalofilové (někteří protisté žijící v pH 10), halofilové (např. řasa Dunaliella salina), barofilové (hlubokomořští sumýši v 110 MPa) i xerofilové (mnohé pouštní houby a lišejníky).[60]
Eukaryota (též Eukarya či česky jaderní) je velká skupina (doména) jednobuněčných a mnohobuněčných organismů, jako jsou například živočichové, rostliny, houby nebo prvoci. Eukaryotická buňka na rozdíl od prokaryotní obsahuje pravé buněčné jádro a množství dalších organel oddělených membránou od okolí. Tyto struktury rozdělují buňku na mnoho menších oddílů (kompartmentů). Eukaryontní organismy mají množství jiných unikátních znaků, jako je například specifická struktura bičíků a řasinek, existence cytoskeletu, určité odchylky ve struktuře genomu (např. DNA rozdělená do jednotlivých chromozomů, přítomnost intronů ve větším množství), schopnost pohlavního rozmnožování a mnohá další metabolická a biochemická specifika.
K eukaryontním organismům se řadí všechny buněčné organismy vyjma bakterií a archeí, tedy prokaryot. Klasifikace eukaryot se v poslední době radikálně mění. Říše rostliny, houby, živočichové a protista se v současnosti nahrazují několika přirozenými skupinami, jako jsou Amoebozoa, Opisthokonta, Rhizaria, Plantae (rostliny), Chromalveolata a Excavata. Odhady týkající se počtu druhů eukaryot se pohybují mezi 5 a 30 miliony, popsáno bylo méně než dva miliony druhů.
Eukaryoter (latin Eucaryota, fra græsk Eucarya, af eu ægte + karyon kerne) er biologiske organismer, hvor cellerne indeholder en eller flere cellekerner[2] – i modsætning til prokaryoter. Udover cellekernen indeholder en eukaryot celle andre organeller, hvor de vigtigste er det endoplasmatiske reticulum, golgiapparatet, cytoskelet, mitokondrier, evt. grønkorn (kloroplaster) og fimrehår (flageller/cilier).
Eukaryoter har en typisk diameter på 10-100 um. Der findes exceptionelle store enkeltcellede eukaryoter, med mange cellekerner, på op til 20 cm i diameter; fx Xenophyophore, som lever på havbunden.[1]
Eukaryoter udgør et af de tre biologiske domæner, mens prokaryoterne udgør de to andre domæner, bakterier og arkæer.
De ældste fossiler fra eukaryoter er 1,8–1,9 milliarder år gamle og er fundet i Kina[3]. Sandsynligvis ligger eukaryoternas oprindelse endnu tidligere, måske så tidligt som for tre milliarder år siden[4].
Der findes flere forskellige hypoteser om, hvordan den eukaryote celle er opstået. I de fleste hypoteser indgår endosymbiose, dvs. at flere forskellige bakterier er gået sammen til at danne en fælles celle, som et fremtrædende element. Når det gælder organellerne mitokondrie og grønkorn (kloroplaster) er endosymbiosen velunderbygget, og i begge tilfælde er det lykkedes at identificere, hvilken bakterie der er optaget i den anden bakterie, hhv. (α-proteobakterier og cyanobakterier). Når det gælder cellekernen, er der ingen alment accepteret hypotese. En mulig oprindelse for cellekernen kunne være, at cellemembranen hos en prokaryot er bugtet indad, indtil den til sidst har omsluttet arvemassen og dannet kernemembranen. Andre prokaryote bakterier med en effektiv energiproduktion og cyanobakterier med fotosyntese er derefter trængt ind i cellen og har trukket lidt af cellemembranen med sig, hvilket forklarer, hvorfor mitokondriet og grønkornet har dobbelt membran. Derefter er mitokondrie, ribosom og andre organeller begyndt at arbejde sammen. Både mitokondrier og grønkorn har egen arvemasse og kopierer sig selvstændigt.
Eukaryoter udviklede meiose, kønnet formering, for cirka 1,2 milliarder år siden, hvorved evolutionen virkelig tog fart.[kilde mangler]
Der har gennem tiden været mange forskellige opdelinger af dette domæne.
Den nuværende polyfyletiske opdeling er: [kilde mangler]
I en alternativ (fylogenetisk) opdeling består eukaryoterne af 2 underdomæner
Eukaryoter (latin Eucaryota, fra græsk Eucarya, af eu ægte + karyon kerne) er biologiske organismer, hvor cellerne indeholder en eller flere cellekerner – i modsætning til prokaryoter. Udover cellekernen indeholder en eukaryot celle andre organeller, hvor de vigtigste er det endoplasmatiske reticulum, golgiapparatet, cytoskelet, mitokondrier, evt. grønkorn (kloroplaster) og fimrehår (flageller/cilier).
Eukaryoter har en typisk diameter på 10-100 um. Der findes exceptionelle store enkeltcellede eukaryoter, med mange cellekerner, på op til 20 cm i diameter; fx Xenophyophore, som lever på havbunden.
Eukaryoter udgør et af de tre biologiske domæner, mens prokaryoterne udgør de to andre domæner, bakterier og arkæer.
Eukaryoten oder Eukaryonten (Eukaryota) (von altgriechisch εὖ eu = ‚richtig‘, ‚gut‘ und κάρυον karyon = ‚Nuss‘, ‚Kern‘) sind eine Domäne der Lebewesen, deren Zellen (Eucyten) einen echten Kern und eine reiche Kompartimentierung haben[1] (Tiere, Pflanzen und Pilze). Hierin unterscheiden sie sich von den beiden übrigen Domänen im System der Lebewesen, den prokaryotischen Bakterien und Archaeen (letztere früher auch Urbakterien genannt), beide mit procytischen Zellen.
Die Zellen der Eukaryoten haben meistens einen Durchmesser von 10 bis 30 µm.[2] Sie sind in der Regel wesentlich größer als die von Prokaryoten, ihr Volumen beträgt etwa das 100- bis 10.000-Fache. Für ein reibungsloses Funktionieren der zellulären Abläufe über größere Entfernungen innerhalb der Zelle sind ein höherer Organisationsgrad und eine Aufteilung des Zellraums in Kompartimente (abgegrenzte Räume) sowie Transport zwischen diesen Kompartimenten günstig. Aus diesem Grund sind eukaryotische Zellen mittels Zellorganellen strukturiert, welche wie die Organe eines Körpers verschiedene Funktionen ausüben. Das namensgebende Organell ist der Zellkern mit dem Hauptanteil des genetischen Materials der eukaryotischen Zellen. Weitere Gene kommen je nach Art in Mitochondrien (Organellen, die durch chemische Reaktionen Energie zur Verfügung stellen), in Einzelfällen auch in den ihnen verwandten Hydrogenosomen sowie in fast allen Plastiden (unter anderem in den Photosynthese betreibenden Chloroplasten) vor. Dem intrazellulären Transport dienen die Organellen des Endomembransystems.
Struktur und Form wird den eukaryotischen Zellen durch das Cytoskelett verliehen, das auch der Fortbewegung dient. Es ist aus Mikrotubuli, Intermediärfilamenten und Mikrofilamenten aufgebaut. Einige Eukaryoten, beispielsweise Pflanzen und Pilze, haben auch Zellwände, die die Zellen außerhalb der Cytoplasmamembran einschließen und ihre Form bestimmen.
Eine weitere Besonderheit der Eukaryoten liegt in der Proteinbiosynthese: Anders als Prokaryoten sind Eukaryoten in der Lage, mittels derselben DNA-Information durch alternatives Spleißen unterschiedliche Proteine herzustellen.
Eukaryoten können Einzeller oder mehrzellige Lebewesen sein. Diese bestehen aus einer größeren Zahl von Zellen mit gemeinsamem Stoffwechsel, wobei spezielle Zelltypen bestimmte Aufgaben übernehmen. Die meisten bekannten Mehrzeller sind Eukaryoten, darunter die Pflanzen, Tiere und mehrzelligen Pilze.
Der Name verweist auf das Vorhandensein eines Zellkerns (Eukaryota/Eukaryonta/Eucarya/Eukarya; zu altgriechisch εὖ eû ‚gut, echt‘ und κάρυον káryon ‚Nuss‘).
In der biologischen Systematik bilden die Eukaryoten eine der drei Domänen, also der höchsten Kategorien zur Klassifizierung von Lebewesen. Die aktuelle Systematik der Eukaryoten wurde von Adl et al. 2019 aufgestellt. Sie gliedert die Eukaryoten wie folgt:[3]
Daneben existieren zahlreiche Taxa mit unsicherer Stellung (incertae sedis), darunter unter anderem
Thomas Cavalier-Smith gliedert die Eukaryoten wie folgt:[5][6][7][6][8][9][10][11]
EukaryotaDie ältesten makroskopischen, mehrzelligen, möglicherweise eukaryotischen Fossilien sind 1,5 Milliarden Jahre alt.[14] Es ist nicht bekannt, ob die hypothetischen evolutionären Vorläufer der Eukaryoten – die sogenannten Urkaryoten[15] – bereits Organellen besaßen (diese wären aufgrund des nachteiligen Oberflächen-Volumen-Verhältnisses nötig gewesen) oder sie erst im Laufe ihrer Evolution hin zu den eigentlichen Eukaryoten erwarben. Die bekannteste Theorie zur Entstehung der Organellen ist die Endosymbiontentheorie, die besagt, dass Mitochondrien und Chloroplasten auf Bakterien zurückgehen, die von den zellbiologisch „primitiven“, räuberisch lebenden Urkaryoten ursprünglich als Nahrung aufgenommen und von denen einige durch Zufall nicht verdaut wurden, sondern eine Symbiose mit dem Urkaryoten eingingen.[16][17]
Der letzte gemeinsame Vorfahr aller heutigen Eukaryoten – in der meist englischen Fachliteratur auch als Last Eukaryotic Common Ancestor (LECA), bezeichnet[18] – sollte sowohl einen abgegrenzten Zellkern als auch Mitochondrien besessen haben, da alle bisher untersuchten Eukaryoten Mitochondrien, mitochondrienartige Organellen oder zumindest Kern-DNA von mitochondrialem Ursprung (übertragen durch lateralen Gentransfer) besitzen. Zwar führte Thomas Cavalier-Smith für rezente amitochondriale Einzeller mit Zellkern, die er für evolutionäre Relikte aus der Zeit vor LECA hielt, die Bezeichnung Archezoa ein,[19] jedoch mehrten sich nachfolgend Hinweise darauf, dass es sich bei den Archezoen um sekundär amitochondriale echte Eukaryoten handelt,[20] das heißt um Abkömmlinge von LECA.
Als Alternative zur Urkaryoten-Hypothese wurde 1998 von Bill Martin und Miklós Müller mit der sog. Wasserstoff-Hypothese (en. hydrogen hypothesis) der Erwerb von Mitochondrien durch Archaeen vorgeschlagen.[21][22] Es hat sich nämlich gezeigt, dass die DNA rezenter (heutiger) Asgard-Archaeen enger mit der DNA in den eukaryotischen Zellkernen verwandt ist, als mit der DNA anderen Archaeen. Die Asgard-Archaeen leben in anoxischen Sedimenten der Ozeane und können mit Bakterien in Symbiose leben. Dies entspricht den Voraussetzungen der Wasserstoff-Hypothese, wonach die ersten eukaryotischen Zellen (Eucyten) durch Erwerb der Mitochondrien unter anoxischen Bedingungen entstanden sind.[23]
Dieser Erwerb könnte vor oder gleichzeitig mit dem des Zellkerns stattgefunden haben und könnte eventuell viralen Ursprungs (unter den NCLDVs) sein („Viral eukaryogenesis“, ein Teil der „Out-of-Virus-Hypothese“).[24] Als Kandidaten werden DNA-Viren wie die ‚Medusaviridae‘ gehandelt;[25] und auch der ‚Pseudomonas Phage 201phi2-1‘[26][27] aus der Gattung Phikzvirus, Familie Myoviridae, ein Riesen-Bakteriophage, bildet bei der Infektion einer Bakterienzelle eine kernähnliche Struktur, die Proteine abhängig von ihrer Funktion trennt,[28] diese kernartige Struktur und ihre Schlüsseleigenschaften wurden auch in den verwandten Phagen gefunden.[29]
Eine im Herbst 2020 veröffentlichte Studie legt anhand von umfangreichen Genomanalysen tatsächlich nahe, dass – obwohl bisher noch keine primär amitochondrialen Eukaryoten gefunden wurden – die Vorfahren der Eukaryonten zuerst ihre komplexes Genom mit den zugehörigen Strukturen, und danach die Mitochondrien (oder Vorläufer davon) erworben haben.[30]
Eine Zusammenfassung dieser Diskussion auf dem Stand von 2019 findet sich bei Traci Watson;[31] zu den unterschiedlichen Standpunkten siehe Guglielmini et al. (2019)[32][33] und Koonin und Yutin (2018).[34]
Nach 12 Jahren Forschungsarbeit berichteten Mikrobiologen 2019, erfolgreich ein sich nur sehr langsam vermehrendes Lokiarchaeon aus Tiefseeschlamm isoliert und kultiviert zu haben.[13][35][36] Candidatus Prometheoarchaeum syntrophicum ist ein, nach Vergleich seines Genoms nahe mit den Eukaryoten verwandtes, extrem langsam wachsendes heterotrophes Archaeon, das Energie aus dem Abbau von Peptiden und Aminosäuren gewinnt und dabei Wasserstoff produziert und abgibt. Das Archaeon gedeiht nur in obligater Syntrophie mit Wasserstoff-metabolierenden Organismen, einer davon ein sulfatreduzierendes Delta-Proeobakterium der Desulfovibrionales. Die Autoren nehmen an, dass treibende Kraft der Endosymbiose, die zu den Eukaryoten führte, ein zweites, in ihrem Modellsystem nicht vorhandenes Bakterium, das künftige Mitochondrium, gewesen wäre, dessen Aufgabe zunächst primär die Beseitigung von für das obligat anaerobe Archaeon giftigen Sauerstoffs gewesen sei. Die Rolle des Mitochondriums im Energiestoffwechsel wäre demnach erst sekundär entstanden. Die Autoren halten eine Schlüsselrolle des Wasserstoff-Metabolismus selbst, wie in alternativen Szenarien der Endosymbiontentheorie[37] vorgeschlagen, für unplausibel. Dementsprechend wäre das Wasserstoff-verwertende Bakterium selbst nicht zum Endosymbionten geworden. Die Hypothese setzt einen frühen Ursprung des Mitochondriums in einer noch primitiven, Archaeen-artigen Wirtszelle voraus und unterscheidet sich darin von alternativen Szenarien wie der „Archezoa“-Hypothese von Thomas Cavalier-Smith, nach denen ein fast fertiger Pro-Eukaryot erst spät ein Mitochondrium erworben hätte. Dementsprechend wäre ein „Einfangen“ des künftigen Mitochondriums über Phagocytose unplausibel, da es an den nötigen internen Strukturen wie einem Cytoskelett gefehlt hätte. Interessanterweise verfügt Prometheoarchaeum aber über tentakel-artige Zellfortsätze, mit denen ein ähnlicher Organismus im Archaikum den hypothetischen Endosymbionten alternativ quasi umschlungen haben könnte.
Die Entstehung der Chloroplasten der Photosynthese betreibenden Eukaryoten durch Endosymbiose stellt einen zeitlich späteren Vorgang dar. Zunächst wurden Cyanobakterien von nicht-phototrophen Eukaryoten aufgenommen, was möglicherweise in dieser Form (primäre Endosymbiose) nur einmal geschah. Die anderen Plastiden (Leukoplasten etc.) leiten sich von diesen ab.
Später entstanden komplexe Plastiden (mit mehr als doppelter Membran und ggf. Nukleosom) durch eine sekundäre Endosymbiose, d. h. (weitere) nicht-phototrophe Eukaryoten nahmen (aus der primären Endosymbiose entstandene) phototrophe Eukaryoten durch Endosymbiose auf (so etwa bei den Apicomplexa[38]).
Die Einteilung von Lebewesen in Prokaryoten und Eukaryoten wurde erstmals von Edouard Chatton für Protisten deutlich herausgestellt und 1925 veröffentlicht.[39][40]
Diese Einteilung trat zunächst mit der Einführung des Drei-Domänen-Systems durch Carl Woese 1977 in den Hintergrund[41][42], eines Ansatzes, der das zelluläre Leben in die drei Domänen Bakterien, Archaeen und Eukaryoten aufteilt.
Im Jahr 1984 wurde von James A. Lake und Kollegen die Eozyten-Hypothese vorgeschlagen: Man hatte entdeckt, dass bei der Form der Ribosomen in der Archaeen-Gruppe der Crenarchaeota (mit der ursprünglichen Bezeichnung Eozyten) und der Eukaryoten eine erstaunliche Ähnlichkeit besteht, dass aber die Form der Ribosomen sowohl bei den Bakterien als auch bei den Euryarchaeota, einer anderen Archaeen-Gruppe, deutlich davon abweicht. Es wurde daher vermutet, dass die Eukaryoten aus den Crenarchaeota hervorgegangen sind.[43][44]
Trotz weiterer Hinweise in den 1980er Jahren bekam die Eozyten-Hypothese erst in den 2000er Jahren mit dem Fortschritt der Genom-Analyse neuen Auftrieb. In einer ganzen Reihe von Archaeen wurden Gene entdeckt, die man in ähnlicher Form auch in Eukaryoten findet. Entsprechend den Ergebnissen einiger Studien wurde in einer Abwandlung statt der Crenarchaeota die diesen nahestehende Gruppe der Thaumarchaeota als Ursprung der Eukaryoten vorgeschlagen. (Die Crenarchaeota und Thaumarchaeota werden zusammen mit einigen anderen Archaeen-Gruppen in der Supergruppe „TACK“ zusammengefasst).[43][45][46][47][48]
Mit dem Aufkommen der Metagenomanalyse ließen sich seit 2015 in Proben aus der Nähe von Hydrothermalquellen Kandidaten für Archaeen-Gruppen identifizieren, die den Eukaryoten noch viel näher stehen müssen, als alle zuvor betrachteten Gruppen. Die erste Gruppe war die der „Lokiarchaeota“, gefunden bei einem hydrothermalen Vent Namens „Loki's Schloss“ im Arktischen Ozean zwischen Mohns- und Knipovitsch-Kamm.[49] Da diese Erkenntnisse lediglich aus einer Metagenom-Analyse stammen und man die betreffenden Mikroben noch bis auf weiteres nicht kultivieren kann, kommt allen fraglichen Taxa lediglich ein 'Kandidatenstatus' zu, gekennzeichnet durch die Anführungszeichen.
Die „Lokiarchaeota“ werden aufgrund von Ähnlichkeit im Genom mit einigen ebenfalls in diesem Zusammenhang jüngst vorgeschlagenen Gruppen („Thorarchaeota“, „Odinarchaeota“ und „Heimdallarchaeota“) zu einer Kandidaten-Gruppe „Asgard“ zusammengefasst, die unter den Archaeen den Eukaryoten somit am nächsten steht und ein Schwester-Taxon zur Supergruppe „TACK“ darstellt.[50][51][52][53]
Um der engen Verwandtschaft der Archaeen mit den Eukaryoten Rechnung zu tragen, hat Thomas Cavalier-Smith sie bereits 2002 in ein gemeinsames Taxon Neomura gestellt, das damit eine Schwestergruppe zu den Bakterien darstellt – allerdings hatte er ursprünglich angenommen, dass die Trennung von Eukaryoten und Archaeen frühzeitig erfolgte, nicht dass die Eukaryoten aus der „Mitte“ der Archaeen entspringen.[54]
Diese neue Einteilung wird unterstützt durch neuere Erkenntnisse, wonach die Verwendung des DNA-Genoms als Träger der Erbinformationen bei Bakterien einerseits und Archaeen und Eukaryoten andererseits unterschiedlichen (möglicherweise viralen: anderer Teil der „Out-of-Virus-Hypothese“) Ursprungs zu sein scheint. Der letzte gemeinsame Vorfahr aller heute bekannten Lebewesen wäre dann ein archaischer zellulärer Organismus der RNA-Welt (mit Ribosomen, d. h. Protein-Synthese) gewesen.[24]
Weitere Unterstützung bekommt die Theorie durch Untersuchung des Aufbaus der membranständigen F- und V-/A-Typ-ATPasen. Die F-Typ-ATPasen der Mitochondrien und Chloroplasten sind mit denen der Bakterien homolog (wie nach der Endosymbiosetheorie zu erwarten). Die V-Typ-ATPasen an Zytoplasma-Membranen (Vakuolen) der Eukaryoten sind zu denen der Archaeen homolog, was eine Abstammung der Eukaryoten aus einem Zweig der Archaeen stützt. Vereinzelte Ausnahmen (F-Typ-ATPasen bei einigen Archaeen-Species[55] und V-Typ-ATPasen bei einigen Bakteriengruppen) werden auf horizontalen Gentransfer zurückgeführt.[56] Zentrale Untereinheiten der ATPasen sind über alle Lebewesen homolog, was für einen LUCA mit einer zumindest primitiven Membranhülle spricht.[57][58]
Eukaryoten oder Eukaryonten (Eukaryota) (von altgriechisch εὖ eu = ‚richtig‘, ‚gut‘ und κάρυον karyon = ‚Nuss‘, ‚Kern‘) sind eine Domäne der Lebewesen, deren Zellen (Eucyten) einen echten Kern und eine reiche Kompartimentierung haben (Tiere, Pflanzen und Pilze). Hierin unterscheiden sie sich von den beiden übrigen Domänen im System der Lebewesen, den prokaryotischen Bakterien und Archaeen (letztere früher auch Urbakterien genannt), beide mit procytischen Zellen.
Chin-hu̍t seng-bu̍t (Eukaryota) sī sè-pau lāi-té ū sè-pau-hu̍t ê seng-bu̍t, pau-koat tōng-bu̍t, si̍t-bu̍t, khún-lūi, kap goân-seng-seng-bu̍t.
Cĭng-hŏk sĕng-ŭk (真核生物) sê dó̤i sá̤-bàu lā̤ ô sá̤-bàu-hŏk gì dăng-sá̤-bàu sĕng-ŭk gâe̤ng dŏ̤-sá̤-bàu sĕng-ŭk gì tūng-chĭng, bău-guák sū-iū gì dông-ŭk, sĭk-ŭk, cĭng-kṳ̄ng gâe̤ng gì-tă ô mŏk-mŏk bău lā̤ gì hók-căk ā-sá̤-bàu giék-gáiu gì sĕng-ŭk.
250px|right|thumb|Ekaryòt Ekaryòt se yon òganis iniselilè (ki gen yon sèl selil) oswa miltiselilè (ki gen plizyè selil), tankou levi, plant ak zannimo; li konpoze avèk selil ekaryotik ki gen ladan yon nwayo kòrèk ak òganèl ki ini relye avèk manbràn nan.
Les eucariotes, c' est des vicantès sacwès ki sont pus avanceyes ki les bactereyes, arkeyobactereyes ey eubactereyes, k' on lome eto procariotes.
Les mwaissès diferinces, c' est les sacwès ki shuvèt, ki les eucariotes ont, et ki les procariotes n' ont nén :
Classicmint les eucariotes estént dispårtis e 4 ringnes: Animalia, Fungi, Plantae et Protista. Po les troes prumîs, on pout aveur on spepieus definixha. Pol cwatrinme, li ringne des protisses, dj' ô bén, çou ki n' est ni biesse, ni plante ni tchampion, gn a co del divize.
So les dierinnès anêyes, gn a yeu des clapantès avanceyes e sistematike et on-z a polou atåvler on meyeu coxhlaedje filodjenetike des eucariotes.
Ene des dierinnès sintezes, e francès, k' a stî fwaite, c' est l' cene propozêye pa Guillaume Lecointre et Hervé Le Guyader dins Classification phylogénétique du vivant (classifiaedje filodjenetike do vicant) k' a vudî ås eplaidreyes Belin.
Les eucariotes, c' est des vicantès sacwès ki sont pus avanceyes ki les bactereyes, arkeyobactereyes ey eubactereyes, k' on lome eto procariotes.
Les mwaissès diferinces, c' est les sacwès ki shuvèt, ki les eucariotes ont, et ki les procariotes n' ont nén :
des ôrganites ki pårtixhèt l' espåce del celule e sacwantès plaeces, avou tchaeke on spepieus ovraedje, come li nawea (ki contént l' ADN), les mitocondreyes, li reticulom indoplasmike, l' aparey da Golgi, les lissozomes, les perocsizomes, les cloroplasses et vacuyoles po les plantes; on citoskelete, bråmint compôzé d' actene et di miyozene; l' ådjeu di fé l' indocitoze. èn ADN, ki s' dispårti ezès cromozomes sol tins do pårtixhaedje del celule. li pårtixhaedje del celule lomêye mitoze ki mete en alaedje les cintriyoles et l' faxhene mitotike. ene prôpe secsuwalité, la k' tchaeke tipe secsuel apoite li mitan do materiå djenetike.Classicmint les eucariotes estént dispårtis e 4 ringnes: Animalia, Fungi, Plantae et Protista. Po les troes prumîs, on pout aveur on spepieus definixha. Pol cwatrinme, li ringne des protisses, dj' ô bén, çou ki n' est ni biesse, ni plante ni tchampion, gn a co del divize.
So les dierinnès anêyes, gn a yeu des clapantès avanceyes e sistematike et on-z a polou atåvler on meyeu coxhlaedje filodjenetike des eucariotes.
Ene des dierinnès sintezes, e francès, k' a stî fwaite, c' est l' cene propozêye pa Guillaume Lecointre et Hervé Le Guyader dins Classification phylogénétique du vivant (classifiaedje filodjenetike do vicant) k' a vudî ås eplaidreyes Belin.
Le eucaryotes es organismos cellular cuje cellulas ha nucleos e altere organellos includite in membranas. Illos forma un dominio (nomine scientific Eukaryota del greco: ευ/eu, bene o vere + κάρυον/karyon, nuce o nucleo), un del tres in que tote le vita cellular es dividite. Illos include le animales, le plantas, le fungos, e altere organismos unicellular e multicellular.
Le eucaryotes es organismos cellular cuje cellulas ha nucleos e altere organellos includite in membranas. Illos forma un dominio (nomine scientific Eukaryota del greco: ευ/eu, bene o vere + κάρυον/karyon, nuce o nucleo), un del tres in que tote le vita cellular es dividite. Illos include le animales, le plantas, le fungos, e altere organismos unicellular e multicellular.
Artikel ieu ngandung bahan-bahan ti Science Primer nu medal ti NCBI, nu, salaku publikasi pamaréntah AS, aya dina domain publik (1).
[
Eukariota (asalé saka basa Yunani "eu" kang tegesé "becik", lan "karyon" kang tegesé nunjuk ing nuklei sel) iku organisme kanthi sel komplèks, ing ngendi bahan-bahan genetikané disusun dadi nuklei kang kaiket membran. Eukariota kalebu kéwan, tetuwuhan, lan jamur—kang akèh-akèhé multiselular—sarta manéka golongan liyané kang diklasifikasikaké kanthi kolèktif minangka protista (akèh ing antarané uniselular). Suwaliké, organisme-organisme liyané, kaya ta bakteri, ora duwé nuklei lan struktur sel komplèks liyané; organisme-organisme kaya iku mau karan prokariota.
Lumrahé, sel eukariota duwé ukuran kang luwih gedhé tinimbang prokariota lan duwé pérangan-pérangan sub-selular kang karan organel. Béda karo prokariota, DNA eukariota disimpen ing kumpulan kromosom kang kasimpen ing njero nuklei kang kabungkus membran nuklei. Saliyané nglakoni panyigaran sel kanthi aseksual, akèh eukariota uga bisa nglakoni réprodhuksi sèksual liwat prosès fusi sel, kang ora tinemu ing prokariota.
Réproduksi eukariota dilakoni liwat sigar sel, kang lumrahé dumadi kanthi mitosis, ya iku prosès sigaré inti sel kang njalari sawijining sel anak nampa dhuplikat utawa salinan saben kromosom kang diduwèni sel indhuk. Ing sakèhé eukariota ana uga réprodhuksi sèksual, ing antarané sel haploid, ya iku sel kang mung duwé siji kromosom saka saben pasang kromosom kang diduwèni sel indhuk kang nglibataké prosès fusi inti sel (singami) lan panyigaran kanthi meiosis kang ngasilaké sel diploid, ya iku sel kang duwé pasangan kromosom kang lengkap.
Eukariota (asalé saka basa Yunani "eu" kang tegesé "becik", lan "karyon" kang tegesé nunjuk ing nuklei sel) iku organisme kanthi sel komplèks, ing ngendi bahan-bahan genetikané disusun dadi nuklei kang kaiket membran. Eukariota kalebu kéwan, tetuwuhan, lan jamur—kang akèh-akèhé multiselular—sarta manéka golongan liyané kang diklasifikasikaké kanthi kolèktif minangka protista (akèh ing antarané uniselular). Suwaliké, organisme-organisme liyané, kaya ta bakteri, ora duwé nuklei lan struktur sel komplèks liyané; organisme-organisme kaya iku mau karan prokariota.
Tu a eukarioten (Eukaryota) uun a biologii hiart ales labenen, wat en selkial hää.
Labenen mä selen, oober saner selkial, hiart tu a prokarioten.
Tu a eukarioten (Eukaryota) uun a biologii hiart ales labenen, wat en selkial hää.
Labenen mä selen, oober saner selkial, hiart tu a prokarioten.
Eukariot vjen nga greqishtja (eu do të thote i vërtetë, karion do të thotë bërthamë) pra flitet për një bërthamë të vërtetë të përmbajtur në një membranë nukleare.
Eukariotët (/ juːˈkærioʊt, -ət /) janë organizmat, qelizat e të cilëve kanë një bërthamë të mbyllur brenda membranave, ndryshe nga prokariotët (Bakteret dhe Arkeat), të cilat nuk kanë organele të lidhura me membranë. [3] [4] [5] Eukaryotes i përkasin domain Eukaryota ose Eukarya. Emri i tyre vjen nga greqishtja εὖ (eu, "mirë" ose "e vërtetë") dhe κάρυον (karyon, "arrë" ose "kernel"). [6] Qelizat eukariote gjithashtu përmbajnë organele të tjera të lidhura me membranë, siç janë mitokondria dhe aparati Golgi, dhe përveç kësaj, disa qeliza bimësh dhe algash përmbajnë kloroplast. Për dallim nga arkeat njëqelizore dhe bakteret, eukariotët gjithashtu mund të jenë shumëqelizorë dhe të përfshijnë organizmat që përbëhen nga shumë lloje qelizash që formojnë lloje të ndryshme indesh. Kafshët dhe bimët janë eukariotët më të njohur.
Eukariotët mund të riprodhojnë si në mënyrë aseksuale përmes mitozës ashtu edhe seksualisht përmes mejozës dhe shkrirjes gamete. Në mitozë, një qelizë ndahet për të prodhuar dy qeliza gjenetikisht identike. Në meiozë, replikimi i ADN-së pasohet nga dy raunde të ndarjes së qelizave për të prodhuar katër qeliza të vajzës haploid. Këto veprojnë si qeliza seksuale (gametet). Gamdo gamete ka vetëm një grup kromozomësh, secila një përzierje unike e palës përkatëse të kromozomeve prindërore që vijnë nga rekombinimi gjenetik gjatë mejozës.
Domeni Eukaryota është monofletik dhe përbën një nga fushat e jetës në sistemin me tre fusha. Dy fushat e tjera, Bakteret dhe Archaea, janë prokariote [7] dhe nuk kanë asnjë nga karakteristikat e mësipërme. Eukariotët përfaqësojnë një pakicë të vogël të të gjitha gjallesave. [8] Sidoqoftë, për shkak të madhësisë së tyre përgjithësisht shumë më të madhe, biomasa e tyre kolektive në të gjithë botën vlerësohet të jetë gati e barabartë me atë të prokariotëve. [8] Eukariotët evoluan afërsisht 1.6–2,1 miliard vjet më parë, gjatë eonit Proterozoik.
Eukarioti (grč. εὖ - eu = pravi + κάρυον - karyon = jezgro) su organizmi odnosno ćelije kod kojih je nasljedni materijal smješten u jedru obavijenom posebnom jedarnon membranom. U eukariotskoj ćeliji razvile su se i brojne ćelijske organele kojih nema kod prokariotskih organizama, među kojima su: endoplazmatski retikulum, Golđijev aparat, lizozomi i dr.[1][2]<
Eukarioti pripadaju taksonu Eukarya ili Eukaryota.[3][4][5] Eukarioti mogu se reproducirati i aseksualno putem mitoze i seksualno putem mejoze i spajanja gameta. U mitozi, od jedne ćelije dijeljenjem nastaju dvije genetički identične ćelije. U mejozi, nakon replikacije DNK slijede dva kruga podjele ćelije za proizvodnju četiri kćeri ćelije, od kojih je svaka sa polovinom broja hromosom[]a u odnosu na originalnu roditeljsku ćeliju (nastaju haploidne ćelije). One djeluju kao spolne ćelije (gameti – svaki gamet ima samo jedan iz svakog para hromosoma, svaki je jedinstvena genetička kombinacija oodgovarajućeg para roditeljskih hromosoma). To je rezultat genetičke rekombinacije tokom mejoze.
Domen Eukaryota je monofiletski i čini jedan od tri domena života. Ostala dva, Bacteria i Archaea, su prokarioti[6] i nemaju pomenuta svojstva. Eukarioti predstavljaju malu manjinu svih živih bića. Međutim, zbog mnogo veće količine, u ukupnoj količini biomase na Zemlji, procjenjuje se da su izjednačeni sa prokariotima.[7] Prvi eukarioti pojavili su se prije približno oko 1,6–2,1 milijarde godina (tokom eona proterozoik).
Eukariot vjen nga greqishtja (eu do të thote i vërtetë, karion do të thotë bërthamë) pra flitet për një bërthamë të vërtetë të përmbajtur në një membranë nukleare.
Eukarioti (grč. εὖ - eu = pravi + κάρυον - karyon = jezgro) su organizmi odnosno ćelije kod kojih je nasljedni materijal smješten u jedru obavijenom posebnom jedarnon membranom. U eukariotskoj ćeliji razvile su se i brojne ćelijske organele kojih nema kod prokariotskih organizama, među kojima su: endoplazmatski retikulum, Golđijev aparat, lizozomi i dr.<
Eukarioti pripadaju taksonu Eukarya ili Eukaryota. Eukarioti mogu se reproducirati i aseksualno putem mitoze i seksualno putem mejoze i spajanja gameta. U mitozi, od jedne ćelije dijeljenjem nastaju dvije genetički identične ćelije. U mejozi, nakon replikacije DNK slijede dva kruga podjele ćelije za proizvodnju četiri kćeri ćelije, od kojih je svaka sa polovinom broja hromosom[]a u odnosu na originalnu roditeljsku ćeliju (nastaju haploidne ćelije). One djeluju kao spolne ćelije (gameti – svaki gamet ima samo jedan iz svakog para hromosoma, svaki je jedinstvena genetička kombinacija oodgovarajućeg para roditeljskih hromosoma). To je rezultat genetičke rekombinacije tokom mejoze.
Domen Eukaryota je monofiletski i čini jedan od tri domena života. Ostala dva, Bacteria i Archaea, su prokarioti i nemaju pomenuta svojstva. Eukarioti predstavljaju malu manjinu svih živih bića. Međutim, zbog mnogo veće količine, u ukupnoj količini biomase na Zemlji, procjenjuje se da su izjednačeni sa prokariotima. Prvi eukarioti pojavili su se prije približno oko 1,6–2,1 milijarde godina (tokom eona proterozoik).
Eukariotlar (yun. yei yaxshi, qaqiqiy, butun va kaguop — yadro) — toʻliq shakllangan, haqiqiy yadroga ega boʻlgan hujayrali organizmlar. E.ga suvoʻtlar, yuksak oʻsimliklar, barcha hayvonlar, zamburugʻlar kiradi. E. DNK si yadrodagi xromosomalarda joylashgan boʻlib, gistonli oqsillar bilan birikkan nukleosomalarni hosil qilishda ishtirok etadi. E.ning hujayralarida membranali organoiddar yaxshi rivojlangan; ayrim organoidlari (mitoxondriyalar va xloroplastlar) da DNK va avtonom oqsil sintezlovchi apparat mavjud.
Eukarioto esas biologiala celulo (od organismo ek tala celuli) di quo celulokerno kontenas genetikala materialo (separeso la DNA di citosolo).
Eukarya, ouch waal Eurkaryote (oet 't Grieks: εὖ = ech, gans; κάρυον = kern;) zeen alle organisme woavan de celle 'ne celkern en 'n intern membraon höbbe. In de taxonomie waere de eukarya gezeen es ein van de drie domeine woa in 't laeve verdeild kin waere. De twee anger domeine beheure allebei toe aan de prokaryote, namelik de Archaea en de Bacteria. 't Groot versjil tösse de Eukarya en Protiste is det de Protiste 'ne “simplere” celboew höbbe woa in 't DNA neet ómsjlaote waert door de celkern mer vriej in 't cytoplasma ligk. Behalve 'ne celkern höbbe de meiste eurkayotische cellen ouch anger organelle zowie mitochondrië, chloroplasten of 'n golgi-apparaat.
En biolochía, Eukarya u Eukaryota ye lo dominio d'os organismos que tienen as celulas con verdaders nuclios. En as luengas latinas, l'adaptación correcta d'o termin ye eucariont u eucarionte. As transliteracions «eucaria» u més asobén «eucariota» fan referencia a las celulas que composan estes organismos: as celulas eucariotas. A etimolochía d'o termin ye «Eu», d'o griego bien u buen, y «Karyon», centro u nuclio; en conchunto se puede interpretar como «Bien nucleyau/da».
O conchunto d'os organismos eucarionts incluye dende os organismos pluricelulars en dó que las celulas son especializadas en fer fayenas concretas y talment les sería imposible de sobrevivir anisoladas, dica las muitas (anque no totas) especies unicelulars. A resta d'altros organismos vivos (totz unicelulars) se cuaternan adintro d'os altros dos dominios: Archaea y Eubacteria.
Pertenixen a lo dominio Eukarya cuatro reinos: Animalia, Plantae, Fungi y Protista. Totz ellos tienen caracteristicas parellanas a nivel molecular (estructura d'os lipidos, proteinas, y chenoma) y tienen antipasaus comuns. Se piensa que las suyas orichens fincan fa bells 2 mil millons d'anyadas, tot y que l'alcuerdo no ye unanime. Fósils previos no s'han puesto claseficar facilment.
As primeras trazas que poderían relacionar-se con os grupos modernos d'eucariontes emprenciparon a apareixer fa uns 800 millons d'anyos, mientres que la mayoría d'os grupos fosils que hu se tienen bien definius no apareixioron dica lo Cambrián.
Os eucarionts s'han quaternau en quatre reinos: Protista, Plantae, Animalia y Fungi, que poderían estar cinco seguntes Cavalier-Smith (Os anteriors más o reino Chromista) u encara muitos más perque bells autors heban considerau que muitos d'os organismos unicelulars habrían d'estar clasificaus a desparte, fendo puyar ixa posible zifra de reinos dica la ventena.
O reino Protista ye lo que en taxonomía se clama un grupo parafiletico. Ixo quier decir que los suyos miembros han estau en muitos casos quaternaus asinas perque simplament no enchascaban mica bien adintro d'altras agrupacions; ye per ixo que que la suya diversidat ye semilar a la diversidat fundamental d'os eucarionts.
En a recient clasificación de Adl et al. (2005) s'evitan os problemas d'a clasificación adintro de reinos sustituyindo-ie estes taxons per un numer de grupos filochenicament emparentaus ta los que no s'aplica garra categoría con finalidat de no tener o problema d'a suya posterior actualización.[1] En esta clasificación, os grans clados serian estes:
Se veiga que las formas ameboides o flachelars no indican pertenenza a garra grupo, como s'heba creyiu enantes, sin de creyar nuevos grupos artificials dende lo punto d'envista evolutivo.
En biolochía, Eukarya u Eukaryota ye lo dominio d'os organismos que tienen as celulas con verdaders nuclios. En as luengas latinas, l'adaptación correcta d'o termin ye eucariont u eucarionte. As transliteracions «eucaria» u més asobén «eucariota» fan referencia a las celulas que composan estes organismos: as celulas eucariotas. A etimolochía d'o termin ye «Eu», d'o griego bien u buen, y «Karyon», centro u nuclio; en conchunto se puede interpretar como «Bien nucleyau/da».
O conchunto d'os organismos eucarionts incluye dende os organismos pluricelulars en dó que las celulas son especializadas en fer fayenas concretas y talment les sería imposible de sobrevivir anisoladas, dica las muitas (anque no totas) especies unicelulars. A resta d'altros organismos vivos (totz unicelulars) se cuaternan adintro d'os altros dos dominios: Archaea y Eubacteria.
Pertenixen a lo dominio Eukarya cuatro reinos: Animalia, Plantae, Fungi y Protista. Totz ellos tienen caracteristicas parellanas a nivel molecular (estructura d'os lipidos, proteinas, y chenoma) y tienen antipasaus comuns. Se piensa que las suyas orichens fincan fa bells 2 mil millons d'anyadas, tot y que l'alcuerdo no ye unanime. Fósils previos no s'han puesto claseficar facilment.
As primeras trazas que poderían relacionar-se con os grupos modernos d'eucariontes emprenciparon a apareixer fa uns 800 millons d'anyos, mientres que la mayoría d'os grupos fosils que hu se tienen bien definius no apareixioron dica lo Cambrián.
Os eucarionts s'han quaternau en quatre reinos: Protista, Plantae, Animalia y Fungi, que poderían estar cinco seguntes Cavalier-Smith (Os anteriors más o reino Chromista) u encara muitos más perque bells autors heban considerau que muitos d'os organismos unicelulars habrían d'estar clasificaus a desparte, fendo puyar ixa posible zifra de reinos dica la ventena.
O reino Protista ye lo que en taxonomía se clama un grupo parafiletico. Ixo quier decir que los suyos miembros han estau en muitos casos quaternaus asinas perque simplament no enchascaban mica bien adintro d'altras agrupacions; ye per ixo que que la suya diversidat ye semilar a la diversidat fundamental d'os eucarionts.
En a recient clasificación de Adl et al. (2005) s'evitan os problemas d'a clasificación adintro de reinos sustituyindo-ie estes taxons per un numer de grupos filochenicament emparentaus ta los que no s'aplica garra categoría con finalidat de no tener o problema d'a suya posterior actualización. En esta clasificación, os grans clados serian estes:
Amoebozoa (qualques amebas u ameboflachelaus) Archaeplastida (incluye Plantae d'entre altros grupos) Chromoalveolata (como lo suyo nombre indica, encluye Alveolata y altros grupos que feban parte d'o reino Chromista proposato por Cavalier-Smith en 1981). Excavata (incluye bells protozous flachelaus, ameboides o ameboflachelaus) Opisthokonta (incluye Fungi y Animalia, d'entre altros) RhizariaSe veiga que las formas ameboides o flachelars no indican pertenenza a garra grupo, como s'heba creyiu enantes, sin de creyar nuevos grupos artificials dende lo punto d'envista evolutivo.
De eukaryootn zyn een van de drie domeinn van de taxonomie. Ze omvattn alle bêestn met e celkern, woa da 't DNA in zit. Vroeger wierdn der mo 2 domeinn ounderscheedn (de eukaryootn en de prokaryootn), in 1990 is Carl Woese ofgekomn met de nieuwe indelienge woarin da Bacteria, Archaea en Eukarya ounderscheedn wierdn.
Eukaryootn zyn per definitie organismn va wien da under celln intern georganiseerd zyn in organelln en e verschillig cytoskelet. Zukke organelln kunn byvoorbêeld de kern zyn, die et mêest opvolt, mor byvoordêeld ook et Golgi-apparoat, endoplasmoatisch reticulum, mitochondriën, chloroplastn of de vacuooln.
Et domein Eukaryota omvat klassiek volgende rykn:
De moderne indelienge volgt volgende taxonomie:
Over 't olgemeen zyn eukaryotische celln vele groter of prokaryootn. Ierounder volgt e korte beschryvieng van de interne structuurn van een eukaryotische celle.
De celkern is omgeevn deur e dubbele membroan, veelal de nucleaire enveloppe genoemd. Tusschen beide laagn zit der 20 à 40 nm plekke. Binn de binnste membroan zit et DNA, tegoare met alle eiwittn da et nodig et lik histoonn of alles die nodig is vo transcriptie. De nucleaire enveloppe volt utêen beist de mitose en meiose.
Der zyn drie soortn endoplasmoatisch reticulum (ER): ruw endoplasmoatisch reticulum (RER), gleddig endoplasmoatisch reticulum (SER, van et Iengels smooth), en sarcoplasmoatisch reticulum die in de spiern zit. Et RER is continu met de buutenste membroan van de nucleaire enveloppe en et zyn ruw uutzicht te dankn an 't feit da der ribosoomn ip zittn die dienn vo et mRNA van de transcriptie in eiwit te vertoaln.
Et SER dient ton mêer vo lipidesynthese, koolhydroatnsynthese en sommige posttranslationele modificoaties an eiwittn.
't Sarcoplasmoatisch reticulum fungeert als Ca2+-reservoir. Da Ca2+ wordt vrygeloatn vo de celle te depolariseern: e proces die essentieel is in spiercontractie.
't Golgi-apparoat dient vooral vo eiwittn te sorteern en vo soms ze te modificeern. Et krygt die eiwittn van et ER deur vesikels: e sôrte van klêne bulletjes met eiwittn en allerhande in, ofgeschermd van de reste van de celle deur e membroan. Dientengevolge loopn et ER en et Golgi-apparoat ni continu i mekoar over.
De endosymbiosetheorie stelt da mitochondriën, chloroplastn en alle organelln ofgeleid van plastiden ofkomstig zyn deur da e voorloper van al de eukaryote celln va vdage een andere bacterie et opgegeetn. In plekke van em te verteern, et 'n em echter wel loatn leven: azo kost 'n mee profiteern van de metabolische functies van die celle. In 't geval van mitochondriën is da ton aërobe respiratie (dus O2 gebruukn vo energie te vast te leggn ounder de vorme van ATP), vo chloroplastn is da licht gebruukn vo de fotosynthese en azo sukers te maakn.
Mitochondriën en chloroplastn en alletwêe e dubbele membroan lik de kern. 't Verschil zit em derin da de binnste membroan serieus ingestulpt is (zie fotto): da zyn de cristae. Mitochondriën komn in oast alle eukaryootn vore.
Sommige vesikels en e gespecialiseerde functie. Lysosoomn breekn byvoorbeeld de inhoud van deur endocytose opgenomn eetn of, beist da peroxisoomn peroxiden ofbreekn. Peroxiden zyn molecuuln die een O-O biendieng en, die styf reactief is.
Een actinefilament is e helix van 2 actinekeetns, oungeveer 7 nm in diameter. Et actineskelet zit mêestol just ounder 't membroanippervlak en is verantwoordelik vo zowel celvorm en -rigiditeit als cytoplasmoatische stromieng, die nodig is vo organeln te verdeeln beist celdelieng en versnelde distributie van molecuuln over hêel de celle (tegoare me myosine). 't Stoat ook in vo uutstulpiengn te maakn in celn, e proces da essentieel is in celmigroatie.
Deze filamentn zyn 8 à 12 nm in brêedte en zyn ook stabieler geboundn. Under functie bestoat der o.a. in vo celvorm en -rigiditeit te beoudn lik et actineskelet, mor ook vo cel-cel en cel-matrix (celbiologie) interacties te stabiliseern. Z'en e belangryke rol by de vorm van sommigte organeln lik de kern.
Vele eukaryootn en e soorte steerte, e flagelle genoemd. Structureel zyn ze compleet verschillig van de flageln van prokaryootn. De kerne dervan bestoat uut microtubuli, en an de boasis zitn ze vast an e centriool, e complex van 9 microtubuli. 't Systêem vo alignerieng van de chromosoomn beist mitose en meiose is analoog doaran: an weerskantn eje e centriool, met doarut 'ountstoand' microtubuli die no de andere centriool toegroein. Z'en e buutndiameter van oungeveer 25 nm, en e binndiameter van oungeveer 15 nm. Microtubuli zyn zelve e polymeer van α- en β-tubuline.
Dierlikke celn en gin celwand, gin chloroplastn, en e klêendere vacuole. Dankzy et gebrek an zuk e celwand is e dierlikke cel flexibel en kat n potentieel andere celn opeetn, e belangryk proces van uus immuunsysteem.
Planteceln verschilln ip volgende maniern van d'andere eukaryootn:
By vele eukaryootn gebeurt voortplantieng seksueel: da wil zeggn da der twêe celln met een olve DNA-inhoud me mekoare versmeltn, vo azo e nieuwe celle (en uutendelik organisme) te maakn. Over 't olgemêen gebeurt et creëern van extra celln echter geweun deur mitose, woadeuren da je (in theorie) twêe identieke dochtercelln krygt.
De eukaryootn zyn een van de drie domeinn van de taxonomie. Ze omvattn alle bêestn met e celkern, woa da 't DNA in zit. Vroeger wierdn der mo 2 domeinn ounderscheedn (de eukaryootn en de prokaryootn), in 1990 is Carl Woese ofgekomn met de nieuwe indelienge woarin da Bacteria, Archaea en Eukarya ounderscheedn wierdn.
'E eucariute (d"o grieco εὖ eu 'vero' e κάρυον káryon 'nucleo') song nu d"e dominie tassonomice, 'o liviello cchiù auto d"a crassifecazzione scientìfeca addò se dividono 'e essere vivente. 'O criterio pe' 'a distinzzione 'e chisto dominio 'e ll'ato, procariute, è 'a presenza 'e nucleo 'nterno ben definito e isulato d"o riesto d"a cellula tramite l'nvolucro nucleare, int'ô chillo è racchiusa 'a maggior parte d"o materiale gennetico, 'o DNA ('na parte è cuntenuta int'ê mitocundre). 'E eucariute song quinne 'e organisme vivente uni- o pluricellulare costituite 'e cellule dotate 'e nucleo, distinte d"e procariute (gruppo parafileteco), 'e cue cellule procariute song prive 'e nucleo ben differenzziato.
Eukaryote (juːˈkærɪɒt or -oʊt), she bioag eh, as ayns ny killaghyn oc, ta'n stoo gienetagh er nyn dastey sthie çheshvean baghtal (yn eukaryon). Ta oayllee cur eukaryota er y phossan. Er lhimmey jeh bacteyryn, ta ooilley organeyn bioag nyn eukaryoteyn.
Cha nel yn aght rheynn killaghyn cheddin ec eukaryoteyn as prokaryoteyn. Ayns eukaryoteyn, ta mynphiobanyn tayrn ny kroymasoomyn dooblit veih y cheilley. Ta daa haase rheynn ayn: mitosis as meiosis.
Son y chooid smoo, ta killaghyn eukaryotagh feer ny smoo na prokaryoteyn. Ta ymmodee far-chrackanyn sthie as troggalyn sthie oc (ny mynoltyn), chammah as ushylagh killag jeant jeh mynphiobanyn, mynsnaieyn as mean-snaieyn. Ta paart scanshoil ec ny snaieyn as mynphiobanyn shoh son reaghey as gurneil cummey y chilley. Ta GND chillaghyn eukaryoteyn rheynnit ayns bartyn linnagh (kroymasoomyn), as rish rheynn çheshveanagh, ta rollian mynphiobanagh rheynn ny bartyn shoh.
Eukaryota (vo oidgriachisch εὖ eu ‚guad‘, ‚echt‘ und καρυωτός karyotos ‚wos wia a Nuss ausschaugt‘) san olle Lebewesn (dazua ghean aa olle Viecha und Pflanzna), vo dena de Zejn an Zejkern hom. De Organismen wo koan Zejkern hom, wern Prokaryota (dazua ghean de Bakterien) gnennt.
Nebm de Bakterien und Archaeen san de Eukaryota oane vo drei Domäna in da Systematik vo de Lebewesn.
De Eukaryota wern moast in sechs „Supagruppen“ einteit:[1]
Eukaryota ni kundi kubwa la viumbehai ambavyo vina seli zenye kiini cha seli na utando wa seli. Mimea, wanyama na fungi wote tunaoona huhesabiwa humo.
Zinahesabiwa kwa jumla kama domeni kati ya viumbehai na domeni nyingine ni bakteria na archaea ambavyo ni viumbe vidogo sana vinavyoonekana kwa hadubini tu.
Eukaryota walio wengi wana seli nyingi lakini hasa protisti na sehemu za fungi zina seli moja tu.
Seli za eukaryota ni kubwa kuliko seli za bakteria na archaea. Ndani ya seli zao kuna oganeli ambazo ni kama viungo vya mwili lakini vina shughuli maalumu ndani ya seli.
Oganeli inayojulikana zaidi ni kiini cha seli inayotunza sehemu kubwa ya DNA yake.
Seli za eukaryota huwa pia na kitu kinachofanana na kiunzi cha mifupa kwenye mwili kinachojengwa kwa vimirija vya protini za pekee ambazo ni gumu kiasi kuliko ektoplasma inayojaza seli.
Kimsingi seli za eukaryota huwa na
Los Eucariòtas (del grèc eu, verai e karuon, nuclèu) amasson quatre grands règnes del monde viu: los animals, los campairòls, las plantas e los protistas. Constituisson un grop fòrça larg d'organismes, uni- e pluricellulars, definits per lor estructura cellulara.Los eucariòtas possedisson, per oposicion als procariòtas (arqueobactèris e eubactèris) :
Un campairòl: Amanita muscaria
,______________________________ Diplomonada | | ,_________________________ Microsporidia __| | | | ,_____________________ Parabasalida | | | |___| | ,____________________ Myxomycota | | | | | |____________________ Eugenozoos |___| | | |_____________________ Naegleria | | | |_____________________ Entamoeba |___| | ,_____________ Acrasiomycota | | | | ,____________ Rhodophyta | | | | | | ,__________ Ciliada | | | | | | |___|_____ Dinoflagellata |___| | | | | |________ Apicomplexa | | | | ,_____ Laberintulida | | | | | |___________ Oomycota |___| | | |________ Xanthophyta |___| | |________ Chrysophyta | | | |_________ Phaeophyta | | | |__________ Diatomea | | ,____________ Plantae | | |___| ,__________ Fungi |___| |_______ Animalia (Animal)
I Eukaryota (dal grech, el voeur dì "ver nucleo") hinn on domini, el pussee compless de la biologia. El gh'ha dent 5 regn: Animalia, Fonsg, Plantae, Protista e Chromista.
Se distinguen dai Prokaryota perchè el nucleo cellular el se ved ben, l'è dividuu dal rest de la cellula e gh'è dent bona part del DNA.
Eukaryote (juːˈkærɪɒt or -oʊt), she bioag eh, as ayns ny killaghyn oc, ta'n stoo gienetagh er nyn dastey sthie çheshvean baghtal (yn eukaryon). Ta oayllee cur eukaryota er y phossan. Er lhimmey jeh bacteyryn, ta ooilley organeyn bioag nyn eukaryoteyn.
Cha nel yn aght rheynn killaghyn cheddin ec eukaryoteyn as prokaryoteyn. Ayns eukaryoteyn, ta mynphiobanyn tayrn ny kroymasoomyn dooblit veih y cheilley. Ta daa haase rheynn ayn: mitosis as meiosis.
Eukaryota ni kundi kubwa la viumbehai ambavyo vina seli zenye kiini cha seli na utando wa seli. Mimea, wanyama na fungi wote tunaoona huhesabiwa humo.
Zinahesabiwa kwa jumla kama domeni kati ya viumbehai na domeni nyingine ni bakteria na archaea ambavyo ni viumbe vidogo sana vinavyoonekana kwa hadubini tu.
Eukaryota walio wengi wana seli nyingi lakini hasa protisti na sehemu za fungi zina seli moja tu.
Los Eucariòtas (del grèc eu, verai e karuon, nuclèu) amasson quatre grands règnes del monde viu: los animals, los campairòls, las plantas e los protistas. Constituisson un grop fòrça larg d'organismes, uni- e pluricellulars, definits per lor estructura cellulara.Los eucariòtas possedisson, per oposicion als procariòtas (arqueobactèris e eubactèris) :
d'organets, que devesisson l'espaci cellular en compartiments especializats, coma: lo nuclèu (que conten l'ADN), las mitocondrias, lo reticulum endoplasmic, l'aparelh de Golgi, los lisosòmas, los peroxisómas, los cloroplasts e las vacuòlas en çò de las plantas ; un citoesquelèt, compausat mai que mai d'actina e de miosina ; la facultat a realizar l'endocitòsi ; un ADN devesit en mantun cromosòmas; una division cellulara apelada mitòsi (amb intervencion dels centriòls e fusèl mitòtic) ; una vertadièra reproduccion sexuada.Eukaryota (vo oidgriachisch εὖ eu ‚guad‘, ‚echt‘ und καρυωτός karyotos ‚wos wia a Nuss ausschaugt‘) san olle Lebewesn (dazua ghean aa olle Viecha und Pflanzna), vo dena de Zejn an Zejkern hom. De Organismen wo koan Zejkern hom, wern Prokaryota (dazua ghean de Bakterien) gnennt.
Nebm de Bakterien und Archaeen san de Eukaryota oane vo drei Domäna in da Systematik vo de Lebewesn.
'E eucariute (d"o grieco εὖ eu 'vero' e κάρυον káryon 'nucleo') song nu d"e dominie tassonomice, 'o liviello cchiù auto d"a crassifecazzione scientìfeca addò se dividono 'e essere vivente. 'O criterio pe' 'a distinzzione 'e chisto dominio 'e ll'ato, procariute, è 'a presenza 'e nucleo 'nterno ben definito e isulato d"o riesto d"a cellula tramite l'nvolucro nucleare, int'ô chillo è racchiusa 'a maggior parte d"o materiale gennetico, 'o DNA ('na parte è cuntenuta int'ê mitocundre). 'E eucariute song quinne 'e organisme vivente uni- o pluricellulare costituite 'e cellule dotate 'e nucleo, distinte d"e procariute (gruppo parafileteco), 'e cue cellule procariute song prive 'e nucleo ben differenzziato.
Deng animal, tanaman, fungus, ampong protist ausan dong eukaryote (IPA: /juːˈkærɪɒt/ o IPA: /-oʊt/), organismu nung nu ding cells makabalangkas la kareng komplikadung istrukturang atsu king kilub da ring membrane. Awsan deng nucleus ing dakeng makabalut kareng membrane a yang pamiyaliwa da reng cell a eukaryote kareng cell a prokaryote. Ing presensia ning nucleus ing babie king lagyu da reng organismung deti: ibat ya iti king katayang Griegung ευ, a mangabaldugang "mayap/tutu", ampo ing κάρυον, "nues" (nut)
Ing pamipitna da reng cell (cell division) kareng eukaryotes aliwa ya kareng organismung alang nucleus (prokaryote). Ing malilyari, mapipitna la reng chromosome kapamilatan ning pamangimut a panibalan da reng microtubule. Adua lang uri deng paralan ning pamipitna. King mitosis, mapipitna ya ing metung a cell at magi yang aduang cell a milulupa gene (genetically identical). King meiosis naman, a kailangan king pamiparakal a sexual, dadalan ya king recombination ing metung a cell a diploid (maki aduang kopia ning balang chromosome, metung ibat king balang pengari) king balang paris da reng chromosome ibat king balang pengari. Kaibat, makataduang marapat ing pamipitna ning cell, nung nu adua lang haploid cell (gametes) deng magbunga. Atin ya mung metung a uri ning chromosome ing balang gamete, a kabangal ning paris ibat king balang pengari.
Balamu monophyletic la reng eukaryote, ania ila ing metung kareng atlung sakup ning bie o domains of life. Detang adua pang aliua, detang bacteria ampong archaea, prokaryote la, at ala lu karela deng kaurian (features) a mebanggit king babo. Nanupata, atin lang pamilupa king aspetu ning biochemistry deng archaea, anya kadane do reng archaea king clade a Neomura.
A eukaryote (/juːˈkæri.oʊt/ yoo-KARR-ee-oht) is ony organism whose cells contain a nucleus an ither organelles enclosed within membranes.
|deadurl= (help)
Ang lahat ng bagay na may buhay (mga hayop, mga halaman, mga halamang-singaw, at mga protista) ay may mga eukaryote (IPA: /juːˈkærɪɒt/ o IPA: /-oʊt/). Ito ang mga selula na organisado at naka-pagsamang may estruktura na nasa loob ng mga membrano nila. Ang membrana ay isang uri ng estruktura na bumabalot sa mga selula at mga organelle nito. Ang katangiang ito ang naghihiwalay sa eukaryote mula sa mga prokaryote. Ang nukleus ng mga eukaryote ang nagbibigay ng pangalan nila.
Ang Eukaryote ay mula sa salitang Griyego na ευ, na ibig sabihin ay "totoo o mabuti" at κάρυον, na ibig sabihin ay "pili". Karamihan sa mga selulang ito ay mayroong iba't ibang mga organelles tulad ng mitokondriya, mga kloroplast at mga katawang Golgi. Mayroon din silang flagella na yari sa mga mikrotubulo. Ang mga mikrotubulong ito ay may 9+2 na pagkaayos.
![]() Ang lathalaing ito na tungkol sa Biyolohiya ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Ang lathalaing ito na tungkol sa Biyolohiya ay isang usbong. Makatutulong ka sa Wikipedia sa nito.
Ang lahat ng bagay na may buhay (mga hayop, mga halaman, mga halamang-singaw, at mga protista) ay may mga eukaryote (IPA: /juːˈkærɪɒt/ o IPA: /-oʊt/). Ito ang mga selula na organisado at naka-pagsamang may estruktura na nasa loob ng mga membrano nila. Ang membrana ay isang uri ng estruktura na bumabalot sa mga selula at mga organelle nito. Ang katangiang ito ang naghihiwalay sa eukaryote mula sa mga prokaryote. Ang nukleus ng mga eukaryote ang nagbibigay ng pangalan nila.
A eukaryote (/juːˈkæri.oʊt/ yoo-KARR-ee-oht) is ony organism whose cells contain a nucleus an ither organelles enclosed within membranes.
Deng animal, tanaman, fungus, ampong protist ausan dong eukaryote (IPA: /juːˈkærɪɒt/ o IPA: /-oʊt/), organismu nung nu ding cells makabalangkas la kareng komplikadung istrukturang atsu king kilub da ring membrane. Awsan deng nucleus ing dakeng makabalut kareng membrane a yang pamiyaliwa da reng cell a eukaryote kareng cell a prokaryote. Ing presensia ning nucleus ing babie king lagyu da reng organismung deti: ibat ya iti king katayang Griegung ευ, a mangabaldugang "mayap/tutu", ampo ing κάρυον, "nues" (nut)
Ing pamipitna da reng cell (cell division) kareng eukaryotes aliwa ya kareng organismung alang nucleus (prokaryote). Ing malilyari, mapipitna la reng chromosome kapamilatan ning pamangimut a panibalan da reng microtubule. Adua lang uri deng paralan ning pamipitna. King mitosis, mapipitna ya ing metung a cell at magi yang aduang cell a milulupa gene (genetically identical). King meiosis naman, a kailangan king pamiparakal a sexual, dadalan ya king recombination ing metung a cell a diploid (maki aduang kopia ning balang chromosome, metung ibat king balang pengari) king balang paris da reng chromosome ibat king balang pengari. Kaibat, makataduang marapat ing pamipitna ning cell, nung nu adua lang haploid cell (gametes) deng magbunga. Atin ya mung metung a uri ning chromosome ing balang gamete, a kabangal ning paris ibat king balang pengari.
Balamu monophyletic la reng eukaryote, ania ila ing metung kareng atlung sakup ning bie o domains of life. Detang adua pang aliua, detang bacteria ampong archaea, prokaryote la, at ala lu karela deng kaurian (features) a mebanggit king babo. Nanupata, atin lang pamilupa king aspetu ning biochemistry deng archaea, anya kadane do reng archaea king clade a Neomura.
Unner Eukaryoten oder Eukaryonten (Eukaryota/Eukaryonta/Eucarya/Eukarya); vun ooldgr. εὖ (eû) „good, echt“ un κάρυον (káryon) „Nööt“ weert all Leevwesen tohopenfaat, de ehre Zellen en Zellkarn hefft. Just dor wiest de Naam up hen. Blangen de Bakterien un de Archaeen sünd de Eukaryoten een vun de dree Domänen in de bioloogsche Systematik. Bakterien un Archaeen weert tosamenfaat unner den Begreep Prokaryoten. De hefft alltohopen keen Zellkarn.
De Eukaryoten ehre Zellen hefft mehrstendeels en Döörmeter vun 10-30 µm.[1] Normolerwiese sünd se dor veel grötter mit, as de Prokaryoten ehre Zellen. Ehr Volumen is 100 bit 10.000 mol so groot. Dormit in de Zellen mit ehre grötteren Afstänne allens leifig aflopen deit, wat dor nödig is, mütt se good organiseert ween. De Zellruum is updeelt in Unnerdeele (afgrenzte Rüüme) un dat gifft Transport twuschen düsse Unnerdeele in de Zellen. Vundeswegen hefft de Zellen so nömmte Organellen rutbillt, de, just so, as de Organe in’t Lief, de Arbeit overnehmen doot. Dat Organell, wat den Naam geven hett, is de Zellkarn. Dor finnt sik de grote Deel vun dat geneetsche Materiol vun de eukaryotschen Zellen in. Annere Gene gifft dat, je nadem, in de Mitochondern-Organellen, de as „Kraftwark“ for de Zellen arbeiden doot. Se stellt dör cheemsche Reaktschoon Energie praat. Ok in de Plastiden kaamt Gene vör. Dat sünd Organellen, de Photosynthese bedrievt. For den Transport in de Zellen sünd de Organellen vun dat Endomembransystem tostännig.
Struktur un Form kriegt de Zellen vun de Eukaryoten dör dat Cytoskelett, wo se sik ok mit rögen könnt. Dat is tosamensett ut Mikrotubuli, Intermediärfilamente un Mikrofilamente. Dat gifft ok Eukaryoten, as Planten un Swämme, de hefft en Zellwand. De slutt de Zellen buten um de Cytoplasmamembran rum in un bestimmt de Form vun de Zell. Bovenhen könnt de Eukaryoten Protobiosynthese bedrieven.
Eukaryoten könnt Eenzellers oder Mehrzellers ween. Mehrzellers bestaht ut en gröttere Tahl vun Zellen mit gemeensom Stoffwessel. Dor overnehmt denn sunnerliche Zelltypen bestimmte Upgaven bi. De meisten bekannten Mehrzellers sünd Eukaryoten, dormank de Planten, Veelzellers (Deerter) un mehrzelligen Swämme.
In de Systematik vun de Biologie billt de Eukaryoten een vun de dree Domänen, de dat gifft. De Domänen sünd de hööchste Kategorie, wo de Leevwesen insorteert weern könnt. De Systematik vun de Eukaryoten, de in’n Momang aktuell is, is 2012 vun Adl et al. tohoopstellt wurrn. Se deelt de Eukaryoten so in:
Denn gifft dat noch en ganze Reeg vun Taxa, bi de dat nich kloor is, wo se henhören deit (incertae sedis), dor höört unner annern to:
De ollsten mehrzelligen, unner Umstänn eukaryootschen Fossilien, de een mit dat blote Ooge sehn kann, sünd bi 1,5 Mrd. Johre oold.[2] Dat is bitherto nich bekannt, ob düsse eersten Eukaryoten al Organellen harrt hefft (an un for sik weer dat vunwegen de Grötte nödig ween), oder ob se de eerst later kregen hefft, nadem se bi en annern Grad vun Organisatschoon anlangt weern. De bekannteste Theorie, wie de Organellen tostanne kamen sünd, is de Endosymbiontentheorie. De seggt, datt Mitochondrien un Chloroplasten sik ut Bakterien entwickelt hefft, de sik in de fröhen Eukaryoten inlagert harrn.[3][4]
Adl, S. M., Simpson, A. G. B., Lane, C. E., Lukeš, J., Bass, D., Bowser, S. S., Brown, M. W., Burki, F., Dunthorn, M., Hampl, V., Heiss, A., Hoppenrath, M., Lara, E., le Gall, L., Lynn, D. H., McManus, H., Mitchell, E. A. D., Mozley-Stanridge, S. E., Parfrey, L. W., Pawlowski, J., Rueckert, S., Shadwick, L., Schoch, C. L., Smirnov, A. and Spiegel, F. W.: The Revised Classification of Eukaryotes. Journal of Eukaryotic Microbiology, 59: 429–514, 2012, PDF Online
D'Eukaryoten ass eent vun den 3 Domäner vun de Liewewiesen. Zu hinnen ziele souwuel d'Eenzeller wéi och de Mënsch. Doduerch kann een erkenne wéi se sech vun den Prokaryoten (Bakterien an Archaeën) ënnerscheeden. D'Eukaryote gi just a véier Räicher agedeelt, well de ganze Rescht der grousser Wëssenschaft bis haut nach net komplett bekannt ass.
Dës Andeelung kënnt dovu wéi sech déi eenzel Organismegruppe mat Energie an Nahrung versuergen, wat Follgen op d'Liewensweis, d'Formen an d'Funktioune vun deene verschiddene Liewewiesen huet.
Eukaryoten - Archaeën - Bakterien
Commons: Eukaryota – Biller, Videoen oder AudiodateienUnter Eukaryote oder Eukaryonte (Eukaryota; vu altgriechisch εὖ eu ‚guet‘, ‚ächt‘ un καρυωτός karyotos ‚nussartig‘, mänkmol nit ganz korräkt as κάρυον karyon ‚Nuss‘ ibersetzt) wäre alli Läbewäse zämmegfasst, wu d Zälle ne Zällchärn hän. D Gruppe vu allne Organisme, wu kai Zällchärn hän, wäre Prokaryote gnännt. Näbe dr Bakterie un dr Archäe stelle d Eukaryote aini vu dr drei Domäne in dr Syschtematik vu dr Läbewäse dar.
D Zälle vu dr Eukaryote hän zmaischt e Durmässer vu 10-30 µm[1]. Si sin in dr Regle vyl greßer wie die vu dr Prokaryote, ihre Volume umfasst eppe s 100 bis 10000fach. Fir s Funktioniere vu dr zelluläre Ablaif iber greßeri Entfärnige innerhalb vu dr Zälle isch e hechere Organisationsgrad, e Ufdailig vum Zällruum in Kompartimänt (abgränzti Ryym) un e Transport zwische däne Kompartimänt ginschtig. Us däm Grund sin eukaryotischi Zälle mit Zällorganälle strukturiert, wu wie d Organ vun eme Lyyb verschideni Funktione uusiebe. Dr Zällchärn mit em Hauptaadail vum genetische Material vu dr eukaryotische Zälle het dr Gruppe dr Name gee. Wyteri Gen chemme je no Art in dr Mitochondrie – Organäll, wu dur chemischi Reaktione Energi z Verfiegig stelle – un Plaschtide – Organälle, wu d Photosynthese dryybe - vor. Em intrazälluläre Transport diene d Organälle vum Endomembransyschtem.
Struktur un Form git dr eukaryotische Zälle s Cytoskelett, wu au fir d Furtbewegig dient. S isch us Mikrotubuli, Intermediärfilamänt un Mikrofilamänt ufböue. E Dail Eukaryote, zem Byschpel Pflanze un Pilz, hän au Zällwände, wu d Zälle usserhalb vu dr Cytoplasmamembran yyschließe un ihri Form bstimme.
E wyteri Bsunderhait vu dr Eukaryote lyt in dr Proteinbiosynthese: Andersch wie d Prokaryote sin Eukaryote in dr Lag, mit dr nämlige DNA-Information dur alternativ Splicing unterschidligi Protein härzstelle.
Eukaryote chenne Aizäller oder mehzälligi Läbewäse syy. Die bstehn zmaischt us ere greßere Zahl vu Zälle mit gmainsamem Stoffwächsel, doderby ibernämme speziälli Zälltype bstimmti Ufgabe. Di maischte bekannte Mehzäller sin Eukaryote, dodrunter d Pflanze, Dierer un mehzälligi Pilz.
D Eukaryote stelle aini vu dr drei Domäne in dr biologische Syschtematik, also di hegscht Kategori zur Klassifizierig vu Läbewäse dar. Di zurzyt aktuäll Syschtematik vu dr Eukaryote isch vu Adl et al. 2005 ufgstellt wore.[2] Si glidere d Eukaryote in sechs Gruppe, vylmol „Supergruppe“ gnännt, statt eme klassische Ranguusdruck[3]:
Näbe däne Gruppe wird bi däne Taxa d Stellig as uusicher aagsääne (incertae sedis) un si chenne nit yygordnet wäre:
Dur neieri Forschige wäre d Ebriacea zue dr Cercozoa innerhalb vu dr Rhizaria gstellt[4] un Stephanopogon zue dr Excavata[5][6].
Unter Eukaryote oder Eukaryonte (Eukaryota; vu altgriechisch εὖ eu ‚guet‘, ‚ächt‘ un καρυωτός karyotos ‚nussartig‘, mänkmol nit ganz korräkt as κάρυον karyon ‚Nuss‘ ibersetzt) wäre alli Läbewäse zämmegfasst, wu d Zälle ne Zällchärn hän. D Gruppe vu allne Organisme, wu kai Zällchärn hän, wäre Prokaryote gnännt. Näbe dr Bakterie un dr Archäe stelle d Eukaryote aini vu dr drei Domäne in dr Syschtematik vu dr Läbewäse dar.
Unner Eukaryoten oder Eukaryonten (Eukaryota/Eukaryonta/Eucarya/Eukarya); vun ooldgr. εὖ (eû) „good, echt“ un κάρυον (káryon) „Nööt“ weert all Leevwesen tohopenfaat, de ehre Zellen en Zellkarn hefft. Just dor wiest de Naam up hen. Blangen de Bakterien un de Archaeen sünd de Eukaryoten een vun de dree Domänen in de bioloogsche Systematik. Bakterien un Archaeen weert tosamenfaat unner den Begreep Prokaryoten. De hefft alltohopen keen Zellkarn.
D'Eukaryoten ass eent vun den 3 Domäner vun de Liewewiesen. Zu hinnen ziele souwuel d'Eenzeller wéi och de Mënsch. Doduerch kann een erkenne wéi se sech vun den Prokaryoten (Bakterien an Archaeën) ënnerscheeden. D'Eukaryote gi just a véier Räicher agedeelt, well de ganze Rescht der grousser Wëssenschaft bis haut nach net komplett bekannt ass.
Dës Andeelung kënnt dovu wéi sech déi eenzel Organismegruppe mat Energie an Nahrung versuergen, wat Follgen op d'Liewensweis, d'Formen an d'Funktioune vun deene verschiddene Liewewiesen huet.
Eukaryoty (Eucaryota; wót starogrichiskego εὖ eu „dobry“, „wopšawdny“ a κάρυον karyon „wórjech“, „jědro“) twórje jadnu z tśich domenow w biologiji. Su žywe byśa z wopšawdnym celowym jědrom, kótarež wopśimjeju genetisku maśiznu we formje desoksyribo-nukleinoweje kisaliny (skrot.: DNA, nimski DNS).
Eukaryoty su zwětšego wó wjelerazne wětše ako prokaryoty (něźi 10-100 raz). Aby gładke funkcioněrowanje celularnych wótběgow nad wětšymi dalokosćami nutśika cele zmóžniło, jo wušy organizaciska měra a rozdźělenje ruma cele do kompartimentow (źělne wótrězki) ale teke transport mjazy toś tymi kompartimentami notne. Toś teje pśicyny dla su eukaryotiske cele z pomocu celowych organelow strukturěrowane, kótarež kaž organy śěła rozdźělne funkcije dopołnjuju. Nejwěcej znata organela jo celowe jědro, z głownym późělom genetiskeje maśizny žywego byśa. Dalšne geny wustupuju pó kuždej družynje w mitochondrijach a plastidach.
Dalšna wósebnosć eukaryotow lažy w proteinowej biosyntezy: Hynac ako prokaryoty su eukaryoty zamóžne, z samskeje DNA-informacije pśez alternatiwny splicing rozdźělne proteiny zgótowaś.
Struktura a forma so eukaryotiskej bańce přez cytoskelet da. Bu mjazy drugim z mikrotubulusow, aktinowych włokninow a mikrofilamentow natwarjony.
Eukaryoty bu tradicionelnje do swětow wěcejcelowych zwěrjetow, rostlinow a gribow ale teke jadnocelowych abo wěcejcelowych protistow rozdźělone. Dokulaž protisty pak žedne monofyletisku kupku njetwórje a druge swěty pópšawem z rozdźělnych protistowych kupkow nastajachu, njejo rozdźělenje do swětow wěcej źaržobna.
Ten cas aktualna Systematika eukaryotow bu wót Adl et al. 2005 wuźěłana.[1] Wóni rozrěduju eukaryotow do šesć kupkow:
Eukaryoty (Eucaryota; wót starogrichiskego εὖ eu „dobry“, „wopšawdny“ a κάρυον karyon „wórjech“, „jědro“) twórje jadnu z tśich domenow w biologiji. Su žywe byśa z wopšawdnym celowym jědrom, kótarež wopśimjeju genetisku maśiznu we formje desoksyribo-nukleinoweje kisaliny (skrot.: DNA, nimski DNS).
Eukaryoty su zwětšego wó wjelerazne wětše ako prokaryoty (něźi 10-100 raz). Aby gładke funkcioněrowanje celularnych wótběgow nad wětšymi dalokosćami nutśika cele zmóžniło, jo wušy organizaciska měra a rozdźělenje ruma cele do kompartimentow (źělne wótrězki) ale teke transport mjazy toś tymi kompartimentami notne. Toś teje pśicyny dla su eukaryotiske cele z pomocu celowych organelow strukturěrowane, kótarež kaž organy śěła rozdźělne funkcije dopołnjuju. Nejwěcej znata organela jo celowe jědro, z głownym późělom genetiskeje maśizny žywego byśa. Dalšne geny wustupuju pó kuždej družynje w mitochondrijach a plastidach.
Dalšna wósebnosć eukaryotow lažy w proteinowej biosyntezy: Hynac ako prokaryoty su eukaryoty zamóžne, z samskeje DNA-informacije pśez alternatiwny splicing rozdźělne proteiny zgótowaś.
Struktura a forma so eukaryotiskej bańce přez cytoskelet da. Bu mjazy drugim z mikrotubulusow, aktinowych włokninow a mikrofilamentow natwarjony.
Με τον ελληνικό, διεθνή σήμερα όρο ευκαρυωτικά, ή ευκαρυώτες, ονομάζονται τα κύτταρα τα οποία έχουν πλήρως σχηματισμένο πυρήνα (αρχαία ελληνικά "κάρυον"), εξ ου και η ονομασία τους, σε αντιδιαστολή με κύτταρα που δεν έχουν σχηματισμένο πυρήνα, τα λεγόμενα προκαρυωτικά κύτταρα.
Από αυτό το είδος κυττάρων αποτελούνται όλοι οι πολυκύτταροι οργανισμοί, όπως τα φυτά και τα ζώα καθώς και ορισμένοι μονοκύτταροι οργανισμοί, όπως πρωτόζωα και φύκη. Τα κύτταρα του ανθρώπου είναι επίσης ευκαρυωτικά.
Βασικά μέρη ενός ευκαρυωτικού κυττάρου είναι α) η πλασματική μεμβράνη, β) το κυτταρόπλασμα και ο κυτταρικός πυρήνας. Εξετάζοντας τη δομή αυτών των κυττάρων, παρατηρείται ότι εξωτερικά περικλείονται από μία μεμβράνη και εσωτερικά ο πυρήνας διαχωρίζεται από το υπόλοιπο κύτταρο πάλι με μία μεμβράνη (πυρηνική μεμβράνη). Ανάμεσα στον πυρήνα και στην εξωτερική μεμβράνη, υπάρχουν οργανίδια, τα οποία είναι υπεύθυνα για τις διάφορες λειτουργίες του κυττάρου, όπως μιτοχόνδρια, λυσόσωμα, ριβόσωμα κ.α.
Τα ευκαρυωτικά κύτταρα εμφανίστηκαν στην εξέλιξη της ζωής πολύ αργότερα από τα προκαρυωτικά, τα οποία είναι απλούστερα στην δομή και δεν έχουν πυρήνα.
Οι φυλογενετικές συγγένειες ανάμεσα στους ευκαρυωτικούς οργανισμούς παρουσιάζονται στο πιο κάτω κλαδόγραμμα (τα ονόματα των κλάδων που περιέχουν, μερικώς ή ολικώς, φωτοσυνθέτοντες οργανισμούς ακολουθούνται από ένα πράσινο τετράγωνο): [1]
Ευκάρυα ΜονόκονταΑμοιβόζωα (Amoebozoa)
Μύκητες (Fungi)
Μεσομυκητόζωα (Mesomycetozoa)
Νηματύλια (Filasterea)
Χοανομόναδα (Choanomonada)
Μετάζωα (Animalia)
? Αποδόζωα (Apusozoa)
Αυλακωτά (Excavata) ▇
Φυτά (Plantae) ή Αρχαιοπλαστίδες (Archaeplastida) ▇
Κρυπτόφυτα (Cryptophyta) ▇
Απτόφυτα (Haptophyta) ▇
Αχυρότριχα (Straminopila) ▇
Ριζωτά (Rhizaria) ▇
Βλεφαριδωτά (Ciliata)
Δινομαστιγωτά (Dinoflagellata) ▇
Ακροσυμπλεγματικά (Apicomplexa)
Με τον ελληνικό, διεθνή σήμερα όρο ευκαρυωτικά, ή ευκαρυώτες, ονομάζονται τα κύτταρα τα οποία έχουν πλήρως σχηματισμένο πυρήνα (αρχαία ελληνικά "κάρυον"), εξ ου και η ονομασία τους, σε αντιδιαστολή με κύτταρα που δεν έχουν σχηματισμένο πυρήνα, τα λεγόμενα προκαρυωτικά κύτταρα.
Еукариотите се организми чии клетки (означени како еуцити) имаат јадро во кое генетскиот материјал е одделен од цитоплазмата. Тоа ги прави посложени од прокариотите, со кои ги споделуваат клеточната мембрана и рибозомите. Освен по присуството на јадро, еуцитите се карактеризираат со најразлични цитоплазматични органели и со цитоскелет. Најголемата и најзабележлива органела на еуцитот е јадрото, со дијаметар од околу 5 μm. Генетскиот материјал во јадрото на еуцитите е организиран во линеарни наместо во кружни ДНК молекули. Јадрото е место каде се одвива репликацијата на ДНК и синтезата на РНК, додека транслацијата на РНК во протеини се одвива во рибозомите на цитоплазмата.
Еукариоти се протистите, растенијата, габите и животните. Името доаѓа од грчките зборови ευ, што значи добро/вистинско, и од κάρυον, што значи лешник или јадро.
Освен јадро, еукариотската клетка содржи различни органели обградени со мембрани во цитоплазмата. Овие органели се компартмани во кои се одвиваат различни метаболички процеси. Еуцитите се генерално многу поголеми од прокариотските клетки. Компартманизацијата овозможена од органелите им дозволува на еуцитите да функционираат ефикасно. Две од овие органели — митохондриите и хлоропластите — играат значајни улоги во енергетскиот метаболизам. Митохондриите, кои можат да се најдат кај речиси сите еукариотски клетки, се места на одвивање на оксидативниот метаболизам и се одговорни за производство на поголемиот дел ATP добиен од разградувањето на органските молекули. Хлоропластите се места каде се одвива фотосинтезата и се наоѓаат само во клетките на растенијата и зелените алги. Лизозомите и пероксизомите се исто така специјализирани метаболички компартмани за дигестија на макромолекулите и за различни оксидативни реакции, соодветно. Многу растителни клетки содржат и големи вакуоли кои имаат многу функции, меѓу кои и дигестија на макромолекули и складирање на екскрети и хранливи состојки.
Поради големината и сложеноста на еуцитите, транспортот на протеини до нивните вистински (точно одредени) дестинации во клетката е одговорна задача. Две органели — ендоплазматичен ретикулум и голџиев систем — се наменети за подредување и транспорт на протеините за секреција, вградување во клеточната мембрана и вградување во лизозомите. Ендоплазматичниот ретикулум е издолжена мрежа на внатреклеточни мембрани кои се протегаат од јадрената мембрана низ цитоплазмата. Тој не само што ги обработува и транспортира протеините, туку учествува и во синтезата на липиди. Од ендоплазматичниот ретикулум, протеините се транспортираат во мали мембрански везикули до голџиевиот систем, каде се врши нивната натамошна обработка и подредување за транспорт до нивните крајни дестинации. Освен улогата на протеински транспортер, голџиевиот систем служи како место за липидна синтеза и (кај растителните клетки) како место за синтеза на полисахаридите кои влегуваат во градбата на клеточниот ѕид.
Еуцитите поседуваат цитоскелет (клеточен скелет), кој е друго ниво на внатрешна организација. Тој е мрежа на протеински филаменти кои се протегаат низ цитоплазмата. Дава структурна поддршка на клетката, со што ја одредува клеточната форма и општата организација на цитоплазмата. Тој исто така е одговорен за движењата на цели клетки (како на пример, контракцијата на мускулните клетки) и за внатреклеточниот транспорт и сместување на органелите и другите градбени делови, вклучувајќи ги и движењата на хромозомите за време на делбата на клетките.
Еукариотите се појавиле пред околу 2,7 билиони години, и тоа по околу 1 до 1,5 билиони години од еволуирањето на прокариотите. Проучувањата на нивните ДНК секвенци укажуваат на тоа дека археите и еубактериите се исто толку различни меѓусебе како и од денешните еукариоти. Затоа, еден многу ран настан во еволуцијата по сè изгледа дека бил појавата на три еволутивни линии кои потекнале од еден заеднички предок (археи, еубактерии и еукариоти). Интересно е тоа што многу архејни гени се послични со тие на еукариотите одошто на еубактериите, што укажува на тоа дека археите и еукариотите споделуваат заедничка линија на еволутивно наследство и се поблизу поврзани меѓусебе отколку со еубактериите. Митохондриите и хлоропластите потекнале од ендосимбиотското здружување на аеробни бактерии со цијанобактерии, соодветно, заедно со предците на еукариотите.
Важен чекор во еволуцијата на еуцитите било создавањето на подклеточни органлеи обвиени со мембрани, што поттикнало развој на сложениот карактер на овие клетки. Се смета дека органелите се добиени како резултат на здружувањето на прокариотските клетки со предокот на еукариотите.
Хипотезата за потеклото на еуцитот од симбиотско здружување со прокариоти (ендосимбиоза) е прилично добро поддржана од страна на истражувањата на митохондриите и хлоропластите, за кои се мисли дека потекнале од бактерии што живееле во големи клетки. Овие две органели се слични на бактериите по својата големина, и како и бактериите, се делат со бинарна делба. Исто така, и двете содржат своја сопствена ДНК, која ги кодира некои од нивните составни делови. Митохондриската и хлоропластната ДНК се реплицира секој пат кога ќе се подели органелата, а гените кои таа ги кодира се транскрибираат во органелата и се транслираат до органелните рибозоми. Според ова, митохондриите и хлоропластите содржат нивни сопствени генетски системи, кои се различни од јадрениот геном на клетката. Понатаму, рибозомите и рибозомската РНК на овие органели се поблиски до тие на бактериите отколку на оние кодирани од јадрените геноми на еукариотите.
Денес е општо прифатено ендосимбиотското потекло на овие органели, со што за митхондриите се смета дека еволуирале од аеробни бактерии, а за хлоропластите од фотосинтетски бактерии, какви што се цијанобактериите. Вградувањето на аеробни бактерии им овозможило на анаеробните клетки одвивање на оксидативен метаболизам. Вградувањето на фотосинтетски бактерии дало хранлива независност со развивање на способност за фотосинтеза. Со времето, многу од гените првично застапени кај овие бактерии се вметнале во јадрениот геном на клетката, така што само неколку градбени делови на митохондриите и хлоропластите сè уште се кодирани од органелните геноми.
Еукариотите се организми чии клетки (означени како еуцити) имаат јадро во кое генетскиот материјал е одделен од цитоплазмата. Тоа ги прави посложени од прокариотите, со кои ги споделуваат клеточната мембрана и рибозомите. Освен по присуството на јадро, еуцитите се карактеризираат со најразлични цитоплазматични органели и со цитоскелет. Најголемата и најзабележлива органела на еуцитот е јадрото, со дијаметар од околу 5 μm. Генетскиот материјал во јадрото на еуцитите е организиран во линеарни наместо во кружни ДНК молекули. Јадрото е место каде се одвива репликацијата на ДНК и синтезата на РНК, додека транслацијата на РНК во протеини се одвива во рибозомите на цитоплазмата.
Еукариоти се протистите, растенијата, габите и животните. Името доаѓа од грчките зборови ευ, што значи добро/вистинско, и од κάρυον, што значи лешник или јадро.
Еукарио́ты, (лат.) Eukaryota од (гр.) εὖ еу ‘добрый’ + κάρυον карион ‘ядро’, або ядро́вы,[1]— домена живых организмох,которых обсягують ядро.
Ку еукариотам належать царства:
В сучасности научно описаный понад милион еукариотох, а число вшиткых еукариотичных видох ся одгадуе на понад 9 милионы.[2]
Еукарио́ты, (лат.) Eukaryota од (гр.) εὖ еу ‘добрый’ + κάρυον карион ‘ядро’, або ядро́вы,— домена живых организмох,которых обсягують ядро.
Ку еукариотам належать царства:
Протисты (Protoza), Грибы (Fungi), Ростлины (Plantae), Живина (Animalia), Хромисты (Chromista).В сучасности научно описаный понад милион еукариотох, а число вшиткых еукариотичных видох ся одгадуе на понад 9 милионы.
Штруктура бунок На роздѣл од прокариотох бункы еукариотох суть роздѣлены на цитоплазму и одграничене кариолемов ядро. Але ани цитоплазма еукариотох не е тотожна цитоплазмѣ прокариотох, але мать зложену штруктуру з мембранами, котры формують органоиды бункы. Функция ядра Ядро бункы обсягуе генетичный материал, организованый в хромосомы, котры ся можуть роздѣльовати путьом митозы и так розмѣстньовати генетичный материал меджи дочерныма бунками. По роздѣлѣню ядра роздѣляють ся и бункы. Молекуларна основа хромосомох представлена молекулами DNA, асоциованыма з гистонами и другыма бѣлковинами. Про векшину еукариотичных организмох характерный половый процес росплодожаня.Эукариоттор (Eucаryota). Эукариоттордун клеткаларынын өлчөмү 13 мкмге жакын. Клеткасынын ичи жаргактар менен бөлүнгөн. Клетканын ичиндеги үч мүчөсү (органелла) башка протоплазмалардан эки чел кабыгы менен бөлүнгөн: ядро клеткасы, митохондриялар жана пластидалар (акыркы органеллар жалаң гана өсүмдүкгө тиешелүү). Цитоплазмада ар түрдүү органеллалар жайгашкан, булардын көбүн электрондук микроскоп менен көрүүгө болот. Ошону менен бирге, рибосомалар пластиддерде жана митохондрияларда болот. Клеткадагы бардык органеллала матриксте орун алышкан. Эукариоттордо үч түрдүү клеткалар болот: өсүмдүк, козу карын жана жаныбар клеткалары.
Эукариотторго татал түзүлүштөгү бир жана көп клеткалуу жаныбар, өсүмдүк, балыр, козу карындар жана жөнөкөй түзүлүштөгү организмдер кирет. Бул организмдердин клеткасында ядролору толугу менен жайгашышкан. ДНК ядронун ичинде орун алып, гендери иреттүү түрдө тизилип ар түрдүү түзүлүштөгү хромосомалар түзүлөт.
Тиричилик дүйнөсү укмуштуудай ар түрдүү. Азыркы учурда 500 миңден ашык өсүмдүк түрлөрү, 1,5 миллиондон ашык жаныбар түрлөрү, бактериялардын, көк жашыл балырлар жана миңдеген козу карындын миңдеген түрлөрү изилденип аныкталган. Организмдердин түзүлүшүндөгү кубулууларды байкап түшүндүрүү, алардын теориялык жана иш жүзүндө кандайча пайдалуу же зыяндуу экендигин ачуу жалпы биология илиминин негизги максаты.
Бардык эукариоттордун клеткаларында ядролору бар, алардын ядролору жаргактар менен курчалган. Эукариоттор үч дүйнөгө бөлүнөт: татаал түзүлүштөгү өсүмдүк, козу карындар жана жаныбар. Бул дүйнөнүн өкүлдөрү бири-биринен түзүлүшүдө көптөгөн өзгөчөлүктөрү менен айырмаланат. Көпчүлүк өсүмдүктөр автотрофтук азыктанат, ал эми жаныбар жана козу-карындар гетероторфтук жол менен азыктанат. Бирок бардык татаал түзүлүштү үч дүйнөнүн арасын так бөлүү кыйын. Жаныбар клеткалары жакшы көрүнгөн чел кабык менен капталышкан, жаныбар козу карынга, өсүмдүкгө караганда демилгелүү, кыймылдуу келишип, өзүдөрү ургаачысын тамагын издеп табууга жөндөмдүү. Бирок жаныбардын ичинде жылбай, жабышып бир жерде жашагандары да бар, алар митоздук жана мейоздук жол менен көбөйөт. Денесинде камдалган, белен азыгы, углеводдор жаныбарда гликоген түрүндө топтолот.
Эукарио́таш (эрс: Эукариоты, лат: Eukaryota хьадаьннад къ.-элл. εὖ- «дика» яхача дешахи κάρυον «тIум» яхача дешахи; шира дош: эвкариоты), е тIума — тIум йоалла клеткаш а йолаш дийнача оганизмий цIа (тIехдоалаче) да.
Шейола оганизмаш, фусаши археяши ца лоархIаш, тIум йоаллаш да (вирусаши вироидаши иштта эукариоташ санна лархIац, амма бакъда массаволча биолого лархIац царех дийна оганизмаш).
Эукарио́таш (эрс: Эукариоты, лат: Eukaryota хьадаьннад къ.-элл. εὖ- «дика» яхача дешахи κάρυον «тIум» яхача дешахи; шира дош: эвкариоты), е тIума — тIум йоалла клеткаш а йолаш дийнача оганизмий цIа (тIехдоалаче) да.
Шейола оганизмаш, фусаши археяши ца лоархIаш, тIум йоаллаш да (вирусаши вироидаши иштта эукариоташ санна лархIац, амма бакъда массаволча биолого лархIац царех дийна оганизмаш).
Эукариотлар яки төшлеләр (лат. Eucaryota) — тере организмнарның бер өспатшалыгы. Эукариотларның төп үзенчәлеге — күзәнәк составында төш булу.
Төшлеләргә бактериялардан һәм архейлардан кала барлык организмнар керә.
Үсемлек күзәнәкләрендә шулай ук хлорофилл бөртекләре дә бар.
Эукариотлар яки төшлеләр (лат. Eucaryota) — тере организмнарның бер өспатшалыгы. Эукариотларның төп үзенчәлеге — күзәнәк составында төш булу.
Төшлеләргә бактериялардан һәм архейлардан кала барлык организмнар керә.
Эукарио́ттар (арх. эвкарио́ттар; лат. Eukaryota «яҡшы» + «ядро»), йәки я́дролылар — күҙәнәктәрендә ядролары булған тере организмдарҙың домены (өҫбатшалыҡ). Бактерияларҙан һәм архейҙарҙан башҡалары барыһы ла эукариоттар.

Хайуандар, үҫемлектәр, бәшмәктәр, шулай уҡ, протистар тигән төркөмдәгеләр — барыһы ла эукариот организмдар. Улар бер күҙәнәклеләр һәм күп күҙәнәклеләр булырға мөмкиндәр.
Әммә бөтәһенең дә төҙөлөш планы оҡшаш. Бөтәһенең дә цитоплазма эсендә мембраналар системаһы ныҡ үҫешкән. Нәҫел материалы цитоплазманан ике ҡатлы мембрана менән айырылған.
Бынан тыш мембраналы төҙөлөшлө органоидтар системаһы барлыҡҡа килгән:
Айырмалары ҙур булыуға ҡарамаҫтан, ядролылар юғары рангҡа ҡараған монофилетик таксон тип һанала.
Киң таралған фараздарҙың береһе булып, эукариоттар 1,6—2,1 млрд йыл элек барлыҡҡа килгән тигән фекер һанала.
Эукарио́ттар (арх. эвкарио́ттар; лат. Eukaryota «яҡшы» + «ядро»), йәки я́дролылар — күҙәнәктәрендә ядролары булған тере организмдарҙың домены (өҫбатшалыҡ). Бактерияларҙан һәм архейҙарҙан башҡалары барыһы ла эукариоттар.
Эске мембрана системаһы һәм уның компоненттары Хайуандар, үҫемлектәр, бәшмәктәр, шулай уҡ, протистар тигән төркөмдәгеләр — барыһы ла эукариот организмдар. Улар бер күҙәнәклеләр һәм күп күҙәнәклеләр булырға мөмкиндәр.
Әммә бөтәһенең дә төҙөлөш планы оҡшаш. Бөтәһенең дә цитоплазма эсендә мембраналар системаһы ныҡ үҫешкән. Нәҫел материалы цитоплазманан ике ҡатлы мембрана менән айырылған.
Бынан тыш мембраналы төҙөлөшлө органоидтар системаһы барлыҡҡа килгән:
ике ҡат мембраналылар: митохондриялар, пластидтар бер мембраналылар:лизосомалар, Гольджи аппараты, эндоплазмаретикулумы (ЭПС).Айырмалары ҙур булыуға ҡарамаҫтан, ядролылар юғары рангҡа ҡараған монофилетик таксон тип һанала.
Эўкарыёты (па-лацінску: Eucaryota, ад стар.-грэц. eu — добра, поўнасцю і karyon — ядро) — надцарства жывых арганізмаў, вузы якіх маюць аформленае ядро з храмасомамі[1] ды іншыя арганоіды, абмежаваныя мэмбранамі. Да эўкарыётаў адносяцца разнастайныя аднавузавыя, каляніяльныя і шматвузачныя арганізмы[2]. Слова «эўкарыёт» паходзіць ад слова karion, якое абазначае «ядро» (арэха). Тэрмін «пракарыёта» абазначае «даядравы», а «эўкарыёта» — «які мае добрае, ці сапраўднае, ядро»[3]. Унутры ядра знаходзяцца складаныя храмасомы, у якіх ДНК зьвязаная зь бялкамі. Храмасомы рэгулярна мітатычна дзеляцца. Жгуцікі і расьнічкі эўкарыёт маюць характэрную (9+2)-структуру зь мікратрубачак. Апошнія таксама сустракаюцца ў цытаплязьме, якая складана разьдзеленая мэмбранамі. У вузах усіх эўкарыёт маюцца арганэлы, у тым ліку мітахондрыі. У эўкарыятычных арганізмаў, асабліва ў расьлін, часта таксама сустракаюцца вакуолі, абкружаныя адзінай мэмбранай, ці тонаплястам. Акрамя таго, шмат якія эўкарыёты характарызуюцца дзьвюмя важнымі рысамі, якія адсутнічаюць у бактэрый: інтэграваная шматвузавасьць і плоцевае размнажэньне. У расьлін протаплясты суседніх вузаў зьвязаныя плязмадэсмамі, якія пранізваюць вузавыя сьценкі. У жывёлаў вузавых сьценак няма і вузы разьдзеленыя пераважна сваімі плязматычнымі мэмбранамі[4]. Эўкарыятычныя вузы звычайна большыя за пракарыятычныя.
Напрыканцы 19 стагодзьдзя біёлягі выдзялялі толькі два царствы жывых арганізмаў — расьліны і жывёлы, прычым грыбы адносіліся да расьлін. Грыбы як і расьліны вядуць нерухомы лад жыцьця, але выкарыстоўваюць у якасьці запаснага рэчыва глікаген (як жывёлы), таму пазьней яны былі выдзеленыя ў асобнае царства.
Паводле адной з сучасных клясыфікацый, надцарства Эўкарыёты ўключае чатыры царствы:
![]() — сховішча мультымэдыйных матэрыялаў
— сховішча мультымэдыйных матэрыялаў
Эўкарыёты (па-лацінску: Eucaryota, ад стар.-грэц. eu — добра, поўнасцю і karyon — ядро) — надцарства жывых арганізмаў, вузы якіх маюць аформленае ядро з храмасомамі ды іншыя арганоіды, абмежаваныя мэмбранамі. Да эўкарыётаў адносяцца разнастайныя аднавузавыя, каляніяльныя і шматвузачныя арганізмы. Слова «эўкарыёт» паходзіць ад слова karion, якое абазначае «ядро» (арэха). Тэрмін «пракарыёта» абазначае «даядравы», а «эўкарыёта» — «які мае добрае, ці сапраўднае, ядро». Унутры ядра знаходзяцца складаныя храмасомы, у якіх ДНК зьвязаная зь бялкамі. Храмасомы рэгулярна мітатычна дзеляцца. Жгуцікі і расьнічкі эўкарыёт маюць характэрную (9+2)-структуру зь мікратрубачак. Апошнія таксама сустракаюцца ў цытаплязьме, якая складана разьдзеленая мэмбранамі. У вузах усіх эўкарыёт маюцца арганэлы, у тым ліку мітахондрыі. У эўкарыятычных арганізмаў, асабліва ў расьлін, часта таксама сустракаюцца вакуолі, абкружаныя адзінай мэмбранай, ці тонаплястам. Акрамя таго, шмат якія эўкарыёты характарызуюцца дзьвюмя важнымі рысамі, якія адсутнічаюць у бактэрый: інтэграваная шматвузавасьць і плоцевае размнажэньне. У расьлін протаплясты суседніх вузаў зьвязаныя плязмадэсмамі, якія пранізваюць вузавыя сьценкі. У жывёлаў вузавых сьценак няма і вузы разьдзеленыя пераважна сваімі плязматычнымі мэмбранамі. Эўкарыятычныя вузы звычайна большыя за пракарыятычныя.
Напрыканцы 19 стагодзьдзя біёлягі выдзялялі толькі два царствы жывых арганізмаў — расьліны і жывёлы, прычым грыбы адносіліся да расьлін. Грыбы як і расьліны вядуць нерухомы лад жыцьця, але выкарыстоўваюць у якасьці запаснага рэчыва глікаген (як жывёлы), таму пазьней яны былі выдзеленыя ў асобнае царства.
Паводле адной з сучасных клясыфікацый, надцарства Эўкарыёты ўключае чатыры царствы:
Пратысты Грыбы Расьліны Жывёлыसुकेंद्रिक या युकेरियोट (eukaryote) एक जीव को कहा जाता है जिसकी कोशिकाओं (सेल) में झिल्लियों में बंद असरल ढाँचे हों। सुकेंद्रिक और अकेंद्रिक (प्रोकेरियोट) कोशिकाओं में सबसे बड़ा अंतर यह होता है कि सुकेंद्रिक कोशिकाओं में एक झिल्ली से घिरा हुआ केन्द्रक (न्यूक्लियस) होता है जिसके अन्दर आनुवंशिक (जेनेटिक) सामान होता है। जीवविज्ञान में सुकेंद्रिक कोशिकाओं वाले जीवों के टैक्सोन को 'सुकेंद्रिक' या 'युकेरियोटी' (eukaryota) कहते हैं।[1]
यूनानी भाषा में 'यु' (ευ, eu) का मतलब 'अच्छा' और 'केरी' (καρυ, kary) का मतलब 'बीज' या (बादाम या अख़रोट की) 'गरी' होता है। युकेरियोट कोशिकाओं में एक स्पष्ट केंद्र (केन्द्रक, यानि न्यूक्लियस) होता है इसलिए उन्हें 'अच्छा बीज' या 'युकेरियोट' कहा जाता है। संस्कृत और यूनानी दोनों हिन्द-यूरोपीय भाषाएँ हैं, इसलिए उनमें बहुत से सजातीय शब्द हैं। यही सजातीयता 'यु-सु' में है।[2]
सुकेंद्रिक या युकेरियोट (eukaryote) एक जीव को कहा जाता है जिसकी कोशिकाओं (सेल) में झिल्लियों में बंद असरल ढाँचे हों। सुकेंद्रिक और अकेंद्रिक (प्रोकेरियोट) कोशिकाओं में सबसे बड़ा अंतर यह होता है कि सुकेंद्रिक कोशिकाओं में एक झिल्ली से घिरा हुआ केन्द्रक (न्यूक्लियस) होता है जिसके अन्दर आनुवंशिक (जेनेटिक) सामान होता है। जीवविज्ञान में सुकेंद्रिक कोशिकाओं वाले जीवों के टैक्सोन को 'सुकेंद्रिक' या 'युकेरियोटी' (eukaryota) कहते हैं।
সংকোষকেন্দ্ৰীয় বা ইউকেৰিয়ট (ইংৰাজী: eukaryote; (/juːˈkæri.oʊt/ বা /juːˈkæriət/ বা yoo-karr-ee-oht বা yoo-karr-ee-ət) বুলিলে সেইবোৰ জীৱক বুজায়, যিবোৰৰ কোষৰ কোষকেন্দ্ৰ আৰু অন্যান্য অঙ্গাণু জৈব পৰ্দাৰ দ্বাৰা আবৃত থাকে।
এই জীৱবোৰ ইউকেৰিয়া বা ইউকেৰিয়টা শ্ৰেণীবিন্যাসৰ অন্তৰ্গত। এই জীৱৰ মূল বৈশিষ্ট্য হল এই যে, ইহঁতৰ কোষ পৰ্দা-বেষ্টিত কোষকেন্দ্ৰ সহ অন্যান্য অঙ্গাণু থাকে, যিয়ে ইহঁতক প্ৰ'কোষকেন্দ্ৰীয় জীৱ পৰা পৃথক কৰে।[2][3][4] এই কোষত কোষকেন্দ্ৰৰ উপস্থিতিৰ কাৰণে এই জীৱবোৰৰ নামকৰণ ইউকেৰিয়ট দিয়া হৈছে, যি গ্ৰীক শব্দ ευ (ইউ) আৰু κάρυον (কেৰিউন) পৰা আহিছে।[5] কোষকেন্দ্ৰৰ ওপৰিও এনে জীৱৰ কোষত থকা অন্য আৱৰণযুক্ত অঙ্গাণুবোৰ হল, মাইট’কণ্ড্ৰিয়া আৰু গল্গি বস্তু। ইয়াৰোপৰি উদ্ভিদত ক্লোৰ'প্লাষ্ট নামৰ এক ধৰণৰ অঙ্গাণু থাকে। সংকোষকেন্দ্ৰীয় জীৱ এককোষী বা বহুকোষী হব পাৰে। বহুকোষী জীৱৰ বিভিন্ন ধৰণৰ কোষৰ দ্বাৰা গঠিত অনেক ধৰণৰ কলা থাকে।
সংকোষকেন্দ্ৰীয় জীৱই মাইট'চিছৰ মাধ্যমেৰে অযৌন প্ৰজনন আৰু মিঅ'চিছৰ মাধ্যমেৰে যৌন প্ৰজনন কৰে। অযৌন প্ৰজননৰ ক্ষেত্ৰত ই কোষ বিভাজন হৈ জিনগতভাৱে অভিন্ন দুটা কোষ উৎপন্ন হয়। যৌন প্ৰজননৰ ক্ষেত্ৰত এটা কোষে ডিএনএ ৰেপ্লিকেছন আৰু তাৰ পিছত দুবাৰ কোষ বিভাজনৰ ফলত চাৰিটা নতুন কোষৰ উৎপন্ন কৰে, যাৰ প্ৰত্যেকৰে মূল কোষৰ ক্ৰম'জমৰ অৰ্ধেক সংখ্যক ক্ৰ'মজম থাকে। এই অপত্য কোষবোৰে জননকোষ হিচাপে কাম কৰে।
সমগ্ৰ জীৱজগতৰ তুলনাত সংকোষকেন্দ্ৰীয় জীৱৰ সংখ্যা তেনেই নগণ্য,[6] আনকি মানবদেহৰ সমস্ত কোষৰ সংখ্যা মানুহৰ অন্ত্ৰত বাসকৰা বেক্টেৰিয়াৰ সংখ্যাতকৈও কম।[7] কিন্তু সংকোষকেন্দ্ৰীয় জীৱৰ অধিকাংশ সদস্যৰ আকৃতি ডাঙৰ হোৱাৰ কাৰণে সমগ্ৰ বিশ্বজুৰি সিহঁতৰ সমষ্টিগত জৈৱভৰ প্ৰ'কোষকেন্দ্ৰী জীৱৰ প্ৰায় সমান বুলিব পাৰি।[8]
সংকোষকেন্দ্ৰীয় কোষবোৰ প্ৰ'কোষকেন্দ্ৰীয় কোষবোৰৰ তুলনাত যথেষ্ট ডাঙৰ। এই কোষত অন্তঃআৱৰণী যুক্ত বিভিন্ন অঙ্গাণু আৰু মাইক্ৰ'টিবিউল, মাইক্ৰ'ফিলামেন্ট আৰু ইণ্টাৰমিডিয়েট ফিলামেন্টৰ দ্বাৰা গঠিত কোষ-কঙ্কাল থাকে। এই কোষৰ ডিএনএ কুণ্ডুলীকৃত অবস্থাত ক্ৰ'ম'জম গঠন কৰে, যিবোৰ কেন্দ্ৰ বিভাজনৰ সময় বিভক্ত হৈ পৃথক হৈ যায়।
সংকোষকেন্দ্ৰীয় কোষবোৰত থকা আৱৰণীসমূহক বৰ্তমান সমষ্টিগত ভাবে অন্তঃআৱৰণী তন্ত্ৰ বোলা হয়।[9] কোষৰ কোষকেন্দ্ৰ নিউক্লিয়াৰ আৱৰণী নামৰ এক অসংখ্য ছিদ্ৰযুক্ত দ্বিস্তৰীয় অন্তঃআৱৰণীৰ দ্বাৰা বেষ্টিত থাকে, যাৰ মাজেদি বিভিন্ন কোষীয় উপাদান কোষকেন্দ্ৰৰ ভিতৰ বা বাহিৰলৈ যাব পাৰে। এই আৱৰণীৰ পৰা বিভিন্ন চেপেটা আৰু নলাকৃতিৰ অংশ এণ্ডোপ্লাজনিক ৰেটিকুলাম গঠন হয়, যি কোষৰ প্ৰ'টিন পৰিবহনত সহায় কৰে। ৰাইব'জম নামৰ অঙ্গাণু এণ্ডোপ্লাজমিক ৰেটিকুলামৰ লগত যুক্ত হৈ প্ৰ'টিন তৈয়াৰ কৰে। উৎপন্ন প্ৰ'টিন এণ্ডোপ্লাজমিক ৰেটিকুলামৰ ভিতৰত প্ৰৱেশ কৰি ভেচিকল বা ক্ষুদ্ৰ থলিৰ মাজেদি পৰিবাহিত হৈ গল্গি বস্তু নামৰ চেপেটা থলিৰ দৰে আকৃতি বিশিষ্ট অঙ্গাণুত জমা হয় আৰু পুনৰ্গঠিত হয়।
বিভিন্ন ধৰণৰ ভেছিকলে বিভিন্ন ধৰণৰ কাম সম্পাদন হয়। যেনে, লাইছ'জমত থকা উৎসেচকে প্ৰ'টিন ভাঙি সৰল অণুত পৰিণত কৰাত সহায় কৰে। পাৰক্সিজম নামৰ ভেছিকলে কোষৰ পক্ষে ক্ষতিকাৰক পাৰক্সাইড ভাঙি পেলাই। কিছুমান প্ৰ'টজ'ৱাৰ কোষত এক্সট্ৰুজম নামৰ সংকোচনশীল ভেছিকল থাকে যিবোৰে কোষৰ অতিৰিক্ত পানী কোষৰ বাহিৰলৈ নিক্ষেপ কৰাত সহায় কৰে। উচ্চ শ্ৰেণীৰ উদ্ভিদৰ কোষৰ অধিকাংশ আয়তন ইয়াত থকা বৃহদাকৃতিৰ ভেকুৱল বা বায়ুমোনাৰ দ্বাৰা পৰিপূৰ্ণ হৈ থাকে যিবোৰে কোষৰ অভিস্ৰৱণ চাপ বজায় ৰখাত সহায় কৰে।
প্ৰায় সমস্ত সংকোষকেন্দ্ৰীয় কোষত মাইট’কণ্ড্ৰিয়া নামৰ এক অঙ্গাণু থাকে। এই অঙ্গাণুবোৰত এডেন'চিন ট্ৰাইফছফেট নামৰ অণু উৎপন্ন হয়, যি কোষৰ শক্তি সৰবৰাহ কৰে।[10] এই অঙ্গাণুবোৰ দ্বি-স্তৰযুক্ত লিপিড অণুৰ দ্বাৰা গঠিত আৱৰণৰ দ্বাৰা বেষ্টিত থাকে যাৰ ভিতৰৰ স্তৰত থকা মাইট’কণ্ড্ৰিয়াৰ ভিতৰত ক্ৰিষ্টি নামৰ ভাঁজ থাকে। মাইট’কণ্ড্ৰিয়াৰ নিজস্ব ডিএনএ থাকে। দেখা যায় যে, প্ৰ'টিঅ'বেক্টেৰিয়া নামৰ এক প্ৰকাৰৰ অন্তঃমিথ'জীৱি প্ৰ'কোষকেন্দ্ৰীক জীৱ সংকোষকেন্দ্ৰীয় কোষত প্ৰৱেশ কৰি মাইট’কণ্ড্ৰিয়া নামৰ অঙ্গাণুত পৰিণত হয়।
উদ্ভিদ আৰু বহু শেলাইৰ কোষত ক্ল'ৰ'প্লাষ্ট নামৰ অঙ্গাণু থাকে, যি ছাইন'বেক্টেৰিয়া নামৰ এক প্ৰকাৰৰ অন্তঃমিথ'জীৱি প্ৰ'কোষকেন্দ্ৰী জীৱ এইবোৰ কোষত প্ৰৱেশ কৰাৰ ফলত গঠিত হয়। এইবোৰ অঙ্গাণুত ক্ল'ৰফিল নামৰ এক ধৰণৰ জৈৱ অণু থাকে যি সালোকসংশ্লেষণ পদ্ধতিৰ দ্বাৰা গ্লুক'জ উৎপন্ন কৰাত সহায় কৰে।
সংকোষকেন্দ্ৰীয় জীৱই মাইট'চিছৰ মাধ্যমেৰে অযৌন প্ৰজনন আৰু মিঅ'চিছৰ মাধ্যমেৰে যৌন প্ৰজনন কৰে। অযৌন প্ৰজননৰ ক্ষেত্ৰত ই কোষ বিভাজন হৈ জিনগতভাৱে অভিন্ন দুটা কোষ উৎপন্ন হয়। যৌন প্ৰজননৰ ক্ষেত্ৰত এটা কোষে ডিএনএ ৰেপ্লিকেছন আৰু তাৰ পিছত দুবাৰ কোষ বিভাজনৰ ফলত চাৰিটা নতুন কোষৰ উৎপন্ন কৰে, যাৰ প্ৰত্যেকৰে মূল কোষৰ ক্ৰম'জমৰ অৰ্ধেক সংখ্যক ক্ৰ'মজম থাকে। এই অপত্য কোষবোৰে জননকোষ হিচাপে কাম কৰে।
যৌন প্ৰজননৰ বিৱৰ্তন প্ৰতি সংকোষকেন্দ্ৰীয় জীৱৰ প্ৰাচীন আৰু মৌলিক বৈশিষ্ট্য পৰিলক্ষিত হয়। ডাক্স আৰু ৰ'জাৰৰ মতে, প্ৰথম দিশৰ সংকোষকেন্দ্ৰীয় জীৱৰ মাজত ঐচ্ছিক প্ৰজননৰ ভূমিকা আছিল।[11] পূৰ্ৱতে অযৌন প্ৰাণীৰূপে চিহ্নিত ট্ৰাইকম'নাছ ভাজাইনালিছ আৰু জিয়াৰ্ডিয়া ইণ্টেষ্টিনালিছৰ দেহত মিঅ'চিছৰ বাবে প্ৰয়োজনীয় জিনৰ অৱস্থিত দেখা পোৱা যায়।[12][13]
এডল[1] আৰু বাৰ্কিয়ে[14] ফাইল'জেনেটিক প্ৰমাণ, জিন স্বাক্ষৰ আৰু অঙ্গসংস্থানগত বৈশিষ্ট্যৰ ভিত্তিত সংকোষকেন্দ্ৰীয় জীৱেৰ জীৱনবৃক্ষ ৰচনা কৰিছিল।
ইউকেৰিয়ট ডায়াফোৰেটিক্সকোনো কোনো বিশ্লেষণত হ্যাক্ৰোবিয়া দলটোক (হ্যাপ্টোফাইটা + ক্ৰায়োফাইটা) আৰ্কিপ্লাস্টিডা মহাদলৰ ওচৰত অন্তৰ্ভুক্ত কৰা দেখা যায়।[15] ইয়াৰোপৰি, বৰ্তমানৰ কিছু গবেষণাৰ মতে হ্যাপ্টোফাইটা আৰু ক্ৰায়োফাইটাই একে মন'ফাইলি গঠন নকৰে।[16] হ্যাপ্টোফাইটা এছএআৰ মহাদলৰ আৰু ক্ৰায়োফাইটা আৰ্কিপ্লাস্টিডা মহাদলৰ অন্তৰ্ভুক্ত বুলি উল্লেখ কৰা হৈছে।[17]
পূৰ্ৱৰ দৰেই, দুটা ফ্লেজেলা আৰু এই ফ্লেজেলা যুক্ত দুই ধৰণৰ পূৰ্বপুৰুষৰৰ ওপৰত ভিত্তি কৰি সংকোষকেন্দ্ৰীয় জীৱক বাইকোন্ট (আৰ্কিপ্লাস্টিডা + এসএআৰ মহাদল + এক্সক্যাভেটা) আৰু ইউনিকোন্ট (অ্যামিবোজোয়া + ওপিস্থোটোকোন্ট) নামৰ দুটা ভাগত বিভক্ত কৰা হৈছে।[15][18][19] ২০১২ চনৰ এক গবেষণাত প্ৰায় একে ধৰণৰ শ্ৰেণীবিভাগ কৰা হৈছিল।[20]
২০১০[21], ২০১৩[22] আৰু ২০১৪ চনত[23] তলত দিয়া ধৰণে সংকোষকেন্দ্ৰীয় জীৱৰ শ্ৰেণীবিভাগ কৰা হৈছিল।
ইউকেৰিয়ট২০০৯ চনত ৰোগোজিনে আৰ্কিঅ'প্লাষ্টিডা আৰু অন্যান্য সংকোষকেন্দ্ৰীয় জীৱৰ মাজত সুস্পষ্ট বিভাজন কৰিছিল।[24]
ইউকেৰিয়ট২০০৯ চনৰ এই গবেষণাপত্ৰৰ পূৰ্ৱৰ সূত্ৰবোৰ বিশ্লেষণ কৰি আৰু নতুনকৈ বিশ্লেষণ কৰি তলৰ শ্ৰেণীবিভাগটো কৰা হয়।[25]
ইউকেৰিয়টসংকোষকেন্দ্ৰীয় বা ইউকেৰিয়ট (ইংৰাজী: eukaryote; (/juːˈkæri.oʊt/ বা /juːˈkæriət/ বা yoo-karr-ee-oht বা yoo-karr-ee-ət) বুলিলে সেইবোৰ জীৱক বুজায়, যিবোৰৰ কোষৰ কোষকেন্দ্ৰ আৰু অন্যান্য অঙ্গাণু জৈব পৰ্দাৰ দ্বাৰা আবৃত থাকে।
এই জীৱবোৰ ইউকেৰিয়া বা ইউকেৰিয়টা শ্ৰেণীবিন্যাসৰ অন্তৰ্গত। এই জীৱৰ মূল বৈশিষ্ট্য হল এই যে, ইহঁতৰ কোষ পৰ্দা-বেষ্টিত কোষকেন্দ্ৰ সহ অন্যান্য অঙ্গাণু থাকে, যিয়ে ইহঁতক প্ৰ'কোষকেন্দ্ৰীয় জীৱ পৰা পৃথক কৰে। এই কোষত কোষকেন্দ্ৰৰ উপস্থিতিৰ কাৰণে এই জীৱবোৰৰ নামকৰণ ইউকেৰিয়ট দিয়া হৈছে, যি গ্ৰীক শব্দ ευ (ইউ) আৰু κάρυον (কেৰিউন) পৰা আহিছে। কোষকেন্দ্ৰৰ ওপৰিও এনে জীৱৰ কোষত থকা অন্য আৱৰণযুক্ত অঙ্গাণুবোৰ হল, মাইট’কণ্ড্ৰিয়া আৰু গল্গি বস্তু। ইয়াৰোপৰি উদ্ভিদত ক্লোৰ'প্লাষ্ট নামৰ এক ধৰণৰ অঙ্গাণু থাকে। সংকোষকেন্দ্ৰীয় জীৱ এককোষী বা বহুকোষী হব পাৰে। বহুকোষী জীৱৰ বিভিন্ন ধৰণৰ কোষৰ দ্বাৰা গঠিত অনেক ধৰণৰ কলা থাকে।
সংকোষকেন্দ্ৰীয় জীৱই মাইট'চিছৰ মাধ্যমেৰে অযৌন প্ৰজনন আৰু মিঅ'চিছৰ মাধ্যমেৰে যৌন প্ৰজনন কৰে। অযৌন প্ৰজননৰ ক্ষেত্ৰত ই কোষ বিভাজন হৈ জিনগতভাৱে অভিন্ন দুটা কোষ উৎপন্ন হয়। যৌন প্ৰজননৰ ক্ষেত্ৰত এটা কোষে ডিএনএ ৰেপ্লিকেছন আৰু তাৰ পিছত দুবাৰ কোষ বিভাজনৰ ফলত চাৰিটা নতুন কোষৰ উৎপন্ন কৰে, যাৰ প্ৰত্যেকৰে মূল কোষৰ ক্ৰম'জমৰ অৰ্ধেক সংখ্যক ক্ৰ'মজম থাকে। এই অপত্য কোষবোৰে জননকোষ হিচাপে কাম কৰে।
সমগ্ৰ জীৱজগতৰ তুলনাত সংকোষকেন্দ্ৰীয় জীৱৰ সংখ্যা তেনেই নগণ্য, আনকি মানবদেহৰ সমস্ত কোষৰ সংখ্যা মানুহৰ অন্ত্ৰত বাসকৰা বেক্টেৰিয়াৰ সংখ্যাতকৈও কম। কিন্তু সংকোষকেন্দ্ৰীয় জীৱৰ অধিকাংশ সদস্যৰ আকৃতি ডাঙৰ হোৱাৰ কাৰণে সমগ্ৰ বিশ্বজুৰি সিহঁতৰ সমষ্টিগত জৈৱভৰ প্ৰ'কোষকেন্দ্ৰী জীৱৰ প্ৰায় সমান বুলিব পাৰি।
ਸੁਕੇਂਦਰੀ ਪ੍ਰਾਣੀ ਜਾਂ ਯੂਕੈਰੀਔਟ ਅਜਿਹਾ ਪ੍ਰਾਣੀ ਹੁੰਦਾ ਹੈ ਜੀਹਦੇ ਕੋਸ਼ਾਣੂ ਵਿਚਲੀ ਨਾਭ ਅਤੇ ਹੋਰ ਢਾਂਚਿਆਂ (ਅੰਗਾਣੂ) ਉੱਤੇ ਇੱਕ ਝਿੱਲੀ ਚੜ੍ਹੀ ਹੋਵੇ। ਇਹਨਾਂ ਦੇ ਕੋਸ਼ਾਣੂਆਂ ਦਾ ਅਕੇਂਦਰੀ ਪ੍ਰਾਣੀਆਂ ਦੇ ਕੋਸ਼ਾਣੂਆਂ ਤੋਂ ਇੱਕ ਫ਼ਰਕ ਹੈ ਕਿ ਇਹਨਾਂ ਦੀਆਂ ਨਾਭਾਂ ਉੱਤੇ ਇੱਕ ਨਾਭਕੀ ਝਿੱਲੀ ਚੜ੍ਹੀ ਹੋਈ ਹੁੰਦੀ ਹੈ ਜਿਸ ਅੰਦਰ ਜੀਵਾਣੂ ਪਦਾਰਥ ਮੌਜੂਦ ਹੁੰਦੇ ਹਨ।[3][4][5] ਬਹੁਤੇ ਸੁਕੇਂਦਰੀ ਜੀਵਾਂ ਵਿੱਚ ਮਾਈਟੋਕਾਂਡਰੀਆ ਅਤੇ ਗੌਲਗੀ ਅਪਰੇਟਸ ਵਰਗੇ ਹੋਰ ਝਿੱਲੀਦਾਰ ਅੰਗਾਣੂ ਹੁੰਦੇ ਹਨ। ਇਹਤੋਂ ਇਲਾਵਾ ਬੂਟਿਆਂ ਅਤੇ ਕਾਈਆਂ ਵਿੱਚ ਕਲੋਰੋਪਲਾਸਟ ਵੀ ਹੁੰਦੇ ਹਨ। ਪ੍ਰੋਟੋਜ਼ੋਆ ਵਰਗੇ ਕਈ ਇੱਕ-ਕੋਸ਼ੀ ਜੀਵ ਵੀ ਸੁਕੇਂਦਰੀ ਹੁੰਦੇ ਹਨ। ਸਾਰੇ ਬਹੁਕੋਸ਼ੀ ਜੀਵ ਸੁਕੇਂਦਰੀ ਹੁੰਦੇ ਹਨ ਜਿਵੇਂ ਕਿ ਪੌਦੇ, ਉੱਲੀਆਂ ਅਤੇ ਪ੍ਰਾਣੀ।
மெய்க்கருவுயிரி (Eukaryote) எனப்படுவது, மென்சவ்வுகளால் சூழப்பட்ட சிக்கலான அமைப்புக்களைக் கொண்ட உயிரணுக்களாலான உயிரினம் ஆகும். இது நிலைக்கருவிலி உயிரினங்களிலிருந்து வேறுபடுவது முக்கியமாக மரபணு அல்லது பாரம்பரியப் பொருளைக் கொண்டிருக்கும் நிலையான கருவையும், அதனை மூடியுள்ள கருமென்சவ்வையும் கொண்டிருப்பதனால் ஆகும்[1][2][3]. அனேகமான மெய்க்கருவுயிரிகள் மென்சவ்வால் மூடப்பட்ட இழைமணிகள், பசுங்கனிகம் அல்லது பச்சைய உருமணிகள், கொல்கி உபகரணங்கள் போன்ற நுண்ணுறுப்புக்களைக் கொண்டிருக்கும். தாவரங்கள், விலங்குகள், பூஞ்சைகள் போன்ற பல்கல உயிரினங்கள் யாவும் பொதுவாக இவ்வகை மெய்க்கருவுயிரிகளேயாகும்.
மெய்க்கருவுயிரி கலமானது நிலைக்கருவிலி கலத்தை விட அளவில் பெரியது. மெய்க்கருவுயிரி கலத்தில் ஓர் கரு (கலத்தின் அனுசேபத் தொழிற்பாடுகளைக் கட்டுப்படுத்தும் அலகு) காணப்படும். எனினும் நிலைக்கருவிலி கலத்தில் கரு மென்சவ்வு அற்ற ஓர் போலியான கரு போன்ற DNA திரள் மாத்திரமே காணப்படும்.அத்தோடு மெய்க்கருவுயிரி கலத்தில் மாத்திரமே நுண்ணுறுப்புகள் காணப்படும்.
மெய்க்கருவுயிரி கலங்களை இலகுவான பயன்பாட்டுக்காக தாவரக் கலம், விலங்குக் கலம் எனப் பிரித்து நோக்கலாம்.
விலங்குக் கலத்தில் பச்சையவுருமணியோ, கலச்சுவரோ காணப்படுவதில்லை. இதில் சிறிய தற்காலிகமான புன்வெற்றிடங்களே இருக்கும். இதில் கலச்சுவர் இல்லாததால் இதனால் எந்த வடிவத்தையும் அடைய முடியும். உதாரணமாக மனித வெண்குருதிக் கலங்கள் ஏனைய நோயை ஏற்படுத்தும் கலங்களை விழுங்க முடியும். மனித உடலில் மாத்திரம் 210க்கும் மேற்பட்ட கலவகைகள் உள்ளன.
தாவரக் கலங்கள் கரு உள்ள கலங்களாகும். எனவே இவை மெய்க்கருவுயிரி கலங்களாகும். இவற்றில் ஒளித்தொகுப்புக்குத் தேவையான பச்சையம் காணப்படுவது ஒரு சிறப்பம்சமாகும். இவற்றில் விலங்குக் கலங்களில் காணப்படாத பல விசேட அமைப்புக்கள் உள்ளன:
மெய்க்கருவுயிரி (Eukaryote) எனப்படுவது, மென்சவ்வுகளால் சூழப்பட்ட சிக்கலான அமைப்புக்களைக் கொண்ட உயிரணுக்களாலான உயிரினம் ஆகும். இது நிலைக்கருவிலி உயிரினங்களிலிருந்து வேறுபடுவது முக்கியமாக மரபணு அல்லது பாரம்பரியப் பொருளைக் கொண்டிருக்கும் நிலையான கருவையும், அதனை மூடியுள்ள கருமென்சவ்வையும் கொண்டிருப்பதனால் ஆகும். அனேகமான மெய்க்கருவுயிரிகள் மென்சவ்வால் மூடப்பட்ட இழைமணிகள், பசுங்கனிகம் அல்லது பச்சைய உருமணிகள், கொல்கி உபகரணங்கள் போன்ற நுண்ணுறுப்புக்களைக் கொண்டிருக்கும். தாவரங்கள், விலங்குகள், பூஞ்சைகள் போன்ற பல்கல உயிரினங்கள் யாவும் பொதுவாக இவ்வகை மெய்க்கருவுயிரிகளேயாகும்.
Chṳ̂n-fu̍t Sâng-vu̍t (真核生物, Eukaryote) he khì se-pâu khí-yû se-pâu-fu̍t ke Tân-se-pâu Sâng-vu̍t lâu Tô-se-pâu Sâng-vu̍t ke chúng-chhṳ̂n, kì pâu-koat só-yû thûng-vu̍t, chhṳ̍t-vu̍t, chṳ̂n-khiûn lâu khì-thâ khí-yû yù mo̍k pâu kwó chho̍k ke fu̍k-chha̍p â-se-pâu kiet-kèu ke sâng-vu̍t. Chṳ̂n-fu̍t Sâng-vu̍t lâu Ngièn-fu̍t Sâng-vu̍t ke kîn-pún-sin khî-phe̍t he chhièn-chá ke se-pâu nui-hàm yû se-pâu-fu̍t, yîn-chhṳ́ yî Chṳ̂n-fu̍t lòi miang-miàng liá yit-lui se-pâu. Hí-tô Chṳ̂n-fu̍t se-pâu chûng hàn hàm yû khì-thâ se-pâu hi, yì lia̍p sien thí, ya̍p liu̍k thí, kô ngì kî thí tén. Yù yî khí-yû se-pâu fu̍t, yîn-chhṳ́ Chṳ̂n-fu̍t se-pâu ke se-pâu fûn-lie̍t ko-chhàng lâu mò se-pâu fu̍t ke ngièn fu̍t sâng-vu̍t ya thai put siông-thùng. Chṳ̂n-fu̍t sâng-vu̍t chhai chin fa sông he tân-ngièn sin ke, tû su̍k-yî sâm vet hì-thúng chûng ke Chṳ̂n-fu̍t sâng-vu̍t vet, nang-ngoi lióng-ke vet vì thùng su̍k-yî ngièn fu̍t sâng-vu̍t ke se khiûn lâu kú khiûn. Than yù yî Chṳ̂n-fu̍t sâng-vu̍t lâu kú khiûn chhai yit-sia sâng fa sin-chṳt fò kî yîn siông-kôan sin sông khí-yû yit-thin siông-sṳ sin, yîn-chhṳ́ yû sṳ̀ ya chiông liá lióng-chá khiung-thùng kûi yî Neomura yên-fa kî. Khô-ho̍k-kâ siông-sin, chhiùng kî yîn chṳn kí lòi-khon, Chṳ̂n-fu̍t sâng-vu̍t he se khiûn lâu kú khiûn ke kî yîn yùng-ha̍p thí, kì he mêu-chúng kú khiûn lâu se khiûn khiung sâng, yi chúng kiet-ha̍p ke sán-vu̍t.
Ẹ̀ùkáríọ́tì tabi ahamo eukarioti je awon ohun elemin[1] ti awon ahamo ara won ni koroonu ninu. Eyi lo ya won soto si pròkáríọ́tì, ti ahamo won ko ni koroonu. [2]
Ẹ̀ùkáríọ́tì tabi ahamo eukarioti je awon ohun elemin ti awon ahamo ara won ni koroonu ninu. Eyi lo ya won soto si pròkáríọ́tì, ti ahamo won ko ni koroonu.
Eukaryota, whose members are known as eukaryotes (/juːˈkærioʊts, -əts/), is a diverse domain of organisms whose cells have a nucleus. All animals, plants, fungi, and many unicellular organisms are eukaryotes. They constitute a major group of living things, along with the two groups of prokaryotes, the Bacteria and the Archaea.
The eukaryotes emerged in the Archaea, possibly within the Asgard archaea. This implies that there are only two domains of life, Bacteria and Archaea, with eukaryotes incorporated among the Archaea. Eukaryotes represent a small minority of the number of organisms, but, due to their generally much larger size, their collective global biomass is about equal to that of prokaryotes. Eukaryotes emerged approximately 2.2 billion years ago, during the Proterozoic eon, likely as flagellated cells. These were created by symbiogenesis between an anaerobic Asgard archaean and an aerobic proteobacterium, which formed the mitochondria. A second episode of symbiogenesis with a cyanobacterium created the plants, with chloroplasts. The oldest-known eukaryote fossils, multicellular planktonic organisms belonging to the Gabonionta, were discovered in Gabon in 2023, dating back to 2.1 billion years ago.[5]
Eukaryotic cells contain membrane-bound organelles such as the nucleus, the endoplasmic reticulum, and the Golgi apparatus. Eukaryotes may be either unicellular or multicellular. In comparison, prokaryotes are typically unicellular. Unicellular eukaryotes are sometimes called protists. Eukaryotes can reproduce both asexually through mitosis and sexually through meiosis and gamete fusion.
Eukaryotes are organisms that range from microscopic single cells, such as picozoans under 3 micrometres across,[6] to animals like the blue whale, weighing up to 190 tonnes and measuring up to 33.6 metres (110 ft) long,[7] or plants like the coast redwood, up to 120 metres (390 ft) tall.[8] Many eukaryotes are unicellular; the informal grouping called protists includes many of these, with some multicellular forms like the giant kelp up to 200 feet (61 m) long.[9] The multicellular eukaryotes include the animals, plants, and fungi, but again, these groups too contain many unicellular species.[10] Eukaryotic cells are typically much larger than those of prokaryotes—the bacteria and the archaea—having a volume of around 10,000 times greater.[11][12] Eukaryotes represent a small minority of the number of organisms, but, as many of them are much larger, their collective global biomass is about equal to that of prokaryotes.[13]

Prokaryotes (small cylindrical cells, bacteria, on left) and a single-celled eukaryote, Paramecium
The eukaryotes are a diverse lineage, consisting mainly of microscopic organisms.[14] Multicellularity in some form has evolved independently at least 25 times within the eukaryotes.[15][16] Complex multicellular organisms, not counting the aggregation of amoebae to form slime molds, have evolved within only six eukaryotic lineages: animals, symbiomycotan fungi, brown algae, red algae, green algae, and land plants.[17] Eukaryotes are grouped by genomic similarities, so that groups often lack visible shared characteristics.[14]
The defining feature of eukaryotes is that their cells have a nucleus. This gives them their name, from the Greek εὖ (eu, "well" or "good") and κάρυον (karyon, "nut" or "kernel", here meaning "nucleus").[18] Eukaryotic cells have a variety of internal membrane-bound structures, called organelles, and a cytoskeleton which defines the cell's organization and shape. The nucleus stores the cell's DNA, which is divided into linear bundles called chromosomes;[19] these are separated into two matching sets by a microtubular spindle during nuclear division, in the distinctively eukaryotic process of mitosis.[20]
Eukaryotes differ from prokaryotes in multiple ways, with unique biochemical pathways such as sterane synthesis.[21] The eukaryotic signature proteins have no homology to proteins in other domains of life, but appear to be universal among eukaryotes. They include the proteins of the cytoskeleton, the complex transcription machinery, the membrane-sorting systems, the nuclear pore, and some enzymes in the biochemical pathways.[22]


Eukaryote cells include a variety of membrane-bound structures, together forming the endomembrane system.[23] Simple compartments, called vesicles and vacuoles, can form by budding off other membranes. Many cells ingest food and other materials through a process of endocytosis, where the outer membrane invaginates and then pinches off to form a vesicle.[24] Some cell products can leave in a vesicle through exocytosis.[25]
The nucleus is surrounded by a double membrane known as the nuclear envelope, with nuclear pores that allow material to move in and out.[26] Various tube- and sheet-like extensions of the nuclear membrane form the endoplasmic reticulum, which is involved in protein transport and maturation. It includes the rough endoplasmic reticulum where ribosomes are attached to synthesize proteins, which enter the interior space or lumen. Subsequently, they generally enter vesicles, which bud off from the smooth endoplasmic reticulum.[27] In most eukaryotes, these protein-carrying vesicles are released and further modified in stacks of flattened vesicles (cisternae), the Golgi apparatus.[28]
Vesicles may be specialized; for instance, lysosomes contain digestive enzymes that break down biomolecules in the cytoplasm.[29]

Mitochondria are organelles found in all but one eukaryote, Monocercomonoides, which has secondarily lost its mitochondria.[30] The mitochondrion is commonly called "the powerhouse of the cell",[31] for its function providing energy by oxidising sugars or fats to produce the energy store ATP.[32][33] Mitochondria have two surrounding membranes, each a phospholipid bi-layer; the inner of which is folded into invaginations called cristae where aerobic respiration takes place.[34]
Mitochondria contain their own DNA, which has close structural similarities to bacterial DNA, and which encodes rRNA and tRNA genes that produce RNA which is closer in structure to bacterial RNA than to eukaryote RNA.[35]
Some eukaryotes, such as the metamonads such as Giardia and Trichomonas, and the amoebozoan Pelomyxa, appear to lack mitochondria, but all have been found to contain mitochondrion-derived organelles, such as hydrogenosomes and mitosomes, and thus have lost their mitochondria secondarily.[30] They obtain energy by enzymatic action on nutrients absorbed from the environment. The metamonad Monocercomonoides has also acquired, by lateral gene transfer, a cytosolic sulfur mobilization system which provides the clusters of iron and sulfur required for protein synthesis. The normal mitochondrial iron-sulfur cluster pathway has been lost secondarily.[30][36]

Plants and various groups of algae have plastids as well as mitochondria. Plastids, like mitochondria, have their own DNA and are developed from endosymbionts, in this case cyanobacteria. They usually take the form of chloroplasts which, like cyanobacteria, contain chlorophyll and produce organic compounds (such as glucose) through photosynthesis. Others are involved in storing food. Although plastids probably had a single origin, not all plastid-containing groups are closely related. Instead, some eukaryotes have obtained them from others through secondary endosymbiosis or ingestion.[37] The capture and sequestering of photosynthetic cells and chloroplasts, kleptoplasty, occurs in many types of modern eukaryotic organisms.[38][39]

Many eukaryotes have long slender motile cytoplasmic projections, called flagella, or multiple shorter structures called cilia. These organelles are variously involved in movement, feeding, and sensation. They are composed mainly of tubulin, and are entirely distinct from prokaryotic flagella. They are supported by a bundle of microtubules arising from a centriole, characteristically arranged as nine doublets surrounding two singlets. Flagella may have hairs (mastigonemes), as in many Stramenopiles. Their interior is continuous with the cell's cytoplasm.[40][41]
Microfilamental structures composed of actin and actin-binding proteins, including α-actinin, fimbrin, and filamin are present in submembranous cortical layers and bundles, as well. Motor proteins of microtubules, dynein and kinesin, and myosin of actin filaments, provide dynamic character of the network.[42][43]
Centrioles are often present even in cells and groups that do not have flagella, but conifers and flowering plants have neither. They generally occur in groups that give rise to various microtubular roots. These form a primary component of the cytoskeletal structure, and are often assembled over the course of several cell divisions, with one flagellum retained from the parent and the other derived from it. Centrioles produce the spindle during nuclear division.[44]
The cells of plants and algae, fungi and most chromalveolates, but not animals, have a cell wall, a layer outside the cell membrane, providing the cell with structural support, protection, and a filtering mechanism. The cell wall also prevents over-expansion when water enters the cell.[45]
The major polysaccharides making up the primary cell wall of land plants are cellulose, hemicellulose, and pectin. The cellulose microfibrils are linked via hemicellulosic tethers to form the cellulose-hemicellulose network, which is embedded in the pectin matrix. The most common hemicellulose in the primary cell wall is xyloglucan.[46]

Eukaryotes have a life cycle that involves sexual reproduction, alternating between a haploid phase, where only one copy of each chromosome is present in each cell and a diploid phase, with two copies of each chromosome in each cell. The diploid phase is formed by fusion of two haploid gametes, such as eggs and spermatozoa, to form a zygote; this may grow into a body, with its cells dividing by mitosis, and at some stage produce haploid gametes through meiosis, a division that reduces the number of chromosomes and creates genetic variability.[47] There is considerable variation in this pattern. Plants have both haploid and diploid multicellular phases.[48] Eukaryotes have lower metabolic rates and longer generation times than prokaryotes, because they are larger and therefore have a smaller surface area to volume ratio.[49]
The evolution of sexual reproduction may be a primordial characteristic of eukaryotes. Based on a phylogenetic analysis, Dacks and Roger have proposed that facultative sex was present in the group's common ancestor.[50] A core set of genes that function in meiosis is present in both Trichomonas vaginalis and Giardia intestinalis, two organisms previously thought to be asexual.[51][52] Since these two species are descendants of lineages that diverged early from the eukaryotic evolutionary tree, core meiotic genes, and hence sex, were likely present in the common ancestor of eukaryotes.[51][52] Species once thought to be asexual, such as Leishmania parasites, have a sexual cycle.[53] Amoebae, previously regarded as asexual, are anciently sexual; present-day asexual groups likely arose recently.[54]
In antiquity, the two lineages of animals and plants were recognized by Aristotle and Theophrastus. The lineages were given the taxonomic rank of Kingdom by Linnaeus in the 18th century. Though he included the fungi with plants with some reservations, it was later realized that they are quite distinct and warrant a separate kingdom.[55] The various single-cell eukaryotes were originally placed with plants or animals when they became known. In 1818, the German biologist Georg A. Goldfuss coined the word protozoa to refer to organisms such as ciliates,[56] and this group was expanded until Ernst Haeckel made it a kingdom encompassing all single-celled eukaryotes, the Protista, in 1866.[57][58][59] The eukaryotes thus came to be seen as four kingdoms:
The protists were at that time thought to be "primitive forms", and thus an evolutionary grade, united by their primitive unicellular nature.[58] The disentanglement of the deep splits in the tree of life only really started with DNA sequencing, leading to a system of domains rather than kingdoms as top level rank being put forward by Carl Woese, Otto Kandler, and Mark Wheelis in 1990, uniting all the eukaryote kingdoms in the domain "Eucarya", stating however that "'eukaryotes' will continue to be an acceptable common synonym".[3][60] In 1996, the evolutionary biologist Lynn Margulis proposed to replace Kingdoms and Domains with "inclusive" names to create a "symbiosis-based phylogeny", giving the description "Eukarya (symbiosis-derived nucleated organisms)".[4]
By 2014, a rough consensus started to emerge from the phylogenomic studies of the previous two decades.[10][61] The majority of eukaryotes can be placed in one of two large clades dubbed Amorphea (similar in composition to the unikont hypothesis) and the Diphoda (formerly bikonts), which includes plants and most algal lineages. A third major grouping, the Excavata, has been abandoned as a formal group as it is paraphyletic.[2] The proposed phylogeny below includes only one group of excavates (Discoba),[62] and incorporates the recent proposal that picozoans are close relatives of rhodophytes.[63] The Provora are a group of microbial predators discovered in 2022.[1] The Metamonada have been hard to place, being sister possibly to Discoba, possibly to Malawimonada.[14]
Eukaryotes Diphoda Diaphoretickes Archaeplastida Rhodophyta (red algae)![]()
![]()
![]()
.jpg)
![]()
Discoba (Excavata) _(cropped).jpg)
![]()
![]()
![]()
![]()

The origin of the eukaryotic cell, also known as eukaryogenesis, is a milestone in the evolution of life, since eukaryotes include all complex cells and almost all multicellular organisms. The last eukaryotic common ancestor (LECA) is the hypothetical origin of all living eukaryotes,[65] and was most likely a biological population.[66] It is believed to have been a protist with a nucleus, at least one centriole and flagellum, facultatively aerobic mitochondria, sex (meiosis and syngamy), a dormant cyst with a cell wall of chitin and/or cellulose and peroxisomes.[67][68][69]
An endosymbiotic union between a motile anaerobic archaean and an aerobic alphaproteobacterium gave rise to eukaryotes, with mitochondria. A second endosymbiosis with a cyanobacterium gave rise to the ancestor of plants, with chloroplasts.[64]
The timing of the origin of eukaryotes is hard to determine; Knoll (2006) suggests they developed as much as 2.2 billion years ago. Some acritarchs are known from at least 1.65 billion years ago, and the possible alga Grypania has been found as far back as 2.1 billion years ago.[70][71] The "problematic"[72] fossil Diskagma has been found in paleosols 2.2 billion years old.[72]

Structures proposed to represent "large colonial organisms" have been found in the black shales of the Palaeoproterozoic Francevillian B Formation, in Gabon, dubbed the "Francevillian biota" dated at 2.1 billion years old.[73][5] However, the status of these structures as fossils is contested, with other authors suggesting that they might represent pseudofossils. The oldest fossils than can unambiguously be assigned to eukaryotes are from the Ruyang Group of China, dating to approximately 1.8-1.6 billion years ago.[74] Fossils that are clearly related to modern groups start appearing an estimated 1.2 billion years ago, in the form of red algae, though recent work suggests the existence of fossilized filamentous algae in the Vindhya basin dating back perhaps to 1.6 to 1.7 billion years ago.[75]
The presence of steranes, eukaryotic-specific biomarkers, in Australian shales previously indicated that eukaryotes were present in these rocks dated at 2.7 billion years old,[21][76] but these Archaean biomarkers have been rebutted as later contaminants.[77] The oldest valid biomarker records are only around 800 million years old.[78] In contrast, a molecular clock analysis suggests the emergence of sterol biosynthesis as early as 2.3 billion years ago.[79] The nature of steranes as eukaryotic biomarkers is further complicated by the production of sterols by some bacteria.[80][81]
Whenever their origins, eukaryotes may not have become ecologically dominant until much later; a massive increase in the zinc composition of marine sediments 800 million years ago has been attributed to the rise of substantial populations of eukaryotes, which preferentially consume and incorporate zinc relative to prokaryotes, approximately a billion years after their origin (at the latest).[82]
Eukaryota, whose members are known as eukaryotes (/juːˈkærioʊts, -əts/), is a diverse domain of organisms whose cells have a nucleus. All animals, plants, fungi, and many unicellular organisms are eukaryotes. They constitute a major group of living things, along with the two groups of prokaryotes, the Bacteria and the Archaea.
The eukaryotes emerged in the Archaea, possibly within the Asgard archaea. This implies that there are only two domains of life, Bacteria and Archaea, with eukaryotes incorporated among the Archaea. Eukaryotes represent a small minority of the number of organisms, but, due to their generally much larger size, their collective global biomass is about equal to that of prokaryotes. Eukaryotes emerged approximately 2.2 billion years ago, during the Proterozoic eon, likely as flagellated cells. These were created by symbiogenesis between an anaerobic Asgard archaean and an aerobic proteobacterium, which formed the mitochondria. A second episode of symbiogenesis with a cyanobacterium created the plants, with chloroplasts. The oldest-known eukaryote fossils, multicellular planktonic organisms belonging to the Gabonionta, were discovered in Gabon in 2023, dating back to 2.1 billion years ago.
Eukaryotic cells contain membrane-bound organelles such as the nucleus, the endoplasmic reticulum, and the Golgi apparatus. Eukaryotes may be either unicellular or multicellular. In comparison, prokaryotes are typically unicellular. Unicellular eukaryotes are sometimes called protists. Eukaryotes can reproduce both asexually through mitosis and sexually through meiosis and gamete fusion.
Eŭkariotoj (science: Eukaryota)[1] estas domanio de organismoj, kies ĉelkerno enhavas genetikan materialon (apartiganta la DNA-n de la citosolo).
Kelkaj el la organeloj de eŭkariotaj ĉeloj, plej rimarkindaj la mitokondrioj kaj la kloroplastoj, havas duoblajn membranojn. Oni vaste opinias ke tiuj estas endosimbiotoj. Ankaŭ ĉelkernoj havas duoblajn membranojn, sed kutime ne estas opiniataj esti endosimbiotoj.
Eŭkariotaj ĉeloj estas pli grandaj ol prokariotaj. La volumena malsameco estas ĉirkaŭ milopa. La eŭkariotaj vivaĵoj estas la plej nove aperintaj, kaj inkluzivas kvar el la kvin "regnoj" de la kutima taksonomio -- (mi ne certas pri tiuj nomoj) protistoj, fungoj, plantoj, kaj animaloj -- kun la tri lastaj evoluinte de la unua.
Arbo
Chromalveolata (Rhizaria, Stramenopile, Alveolata, Haptophyta, Cryptophyta ...)
Archaeplastida (ruĝaj algoj, Verdaj plantoj )
En biología y taxonomía, Eukaryota o Eukarya (del griego: εὖ eu —‘bueno’, ‘bien’, 'verdadero'— y κάρυον karyon —‘nuez’, ‘carozo’, ‘núcleo’—) es el dominio (o imperio) que incluye los organismos formados por células con núcleo verdadero. La castellanización adecuada del término es eucariota o eucarionte.[5] Estos organismos constan de una o más células eucariotas, abarcando desde organismos unicelulares hasta verdaderos pluricelulares en los que las diferentes células se especializan para diferentes tareas y que, en general, no pueden sobrevivir de forma aislada.
Pertenecen al dominio o imperio eucariota los reinos de los animales, plantas y hongos, así como varios grupos incluidos en el parafilético reino Protista. Todos ellos presentan semejanzas a nivel molecular (estructura de los lípidos, proteínas y genoma), comparten un origen común, y principalmente, comparten el plan corporal de los eucariotas, muy diferente al de procariotas.
Con excepción de algunos organismos unicelulares, el ciclo de vida eucariota alterna una fase haplonte y otra diplonte, que se consigue mediante la alternancia de meiosis y fecundación, procesos que dan células haplontes y diplontes respectivamente.
Las células eucariotas son generalmente mucho más grandes que las procariotas y están mucho más compartimentadas. Poseen una gran variedad de membranas con núcleo rodeado de la envoltura nuclear, retículo endoplasmático y aparato de Golgi, además de mecanismos para la gemación y fusión de vesículas, incluida la exocitosis y endocitosis. Estructuras internas llamadas orgánulos se encargan de realizar funciones especializadas dentro de la célula. Presencia de lisosomas, peroxisomas y mitocondrias.
También caracteriza a todos los eucariotas un esqueleto interno o endoesqueleto, en este caso llamado citoesqueleto, formado por dos entramados de proteínas: el sistema de microtúbulos y el sistema contráctil de actina/miosina, que desempeñan un papel importante en la definición de la organización y forma de la célula, en el tráfico intracelular (por ejemplo, los movimientos de vesículas y orgánulos) y en la división celular. El característico flagelo eucariota y sus motores moleculares asociados se encuentran anclados al citoesqueleto.
El ADN de las células eucariotas está contenido en un núcleo celular separado del resto de la célula por una doble membrana permeable. El material genético se divide en varios bloques lineales llamados cromosomas, que son separados por un huso microtubular durante la división nuclear. Los cromosomas contienen histonas, varios replicones, centrómeros y telómeros. Hay un característico ciclo celular con segregación mitótica y reproducción sexual por meiosis. Se incluye un complejo de poros nucleares, transporte trans-membranal de ARN y proteínas a través de la envoltura nuclear, intrones y nuevos patrones de procesamiento del ARN utilizando espliceosomas.
Para una comparación con las características procariotas, véase: Tabla comparativa.
La célula eucariota debe en gran parte su forma y capacidad de movimiento al citoesqueleto, ya que le otorga rigidez y flexibilidad. En los organismos flagelados ancla los flagelos al resto de la célula y permite su batido durante la locomoción o para la creación de corrientes de agua que le lleven el alimento. En los organismos ameboides permite la extensión de "pies" o seudópodos para la locomoción o la alimentación. También fija los surcos de alimentación de los excavados y el complejo apical que permite a los apicomplejos entrar en las células parasitadas.
Solo después de desarrollar su citoesqueleto pudo el eucariota ancestral realizar la fagocitosis, ya que es este el que, mediante crecimiento diferencial de sus fibras, logra que la célula se deforme para que la fagocitosis ocurra. La fagocitosis es también una propiedad ancestral de los eucariotas, si bien se ha perdido en grupos que se adaptaron a otras formas de alimentación. Hongos y plantas perdieron esta capacidad al desarrollar una pared celular rígida externa a la célula, pero ya contaban con otros modos de nutrición, la saprotrofia o el parasitismo en hongos y la fotosíntesis en plantas.
La mitocondria, derivada de la fagocitosis y posterior simbiogénesis de una proteobacteria, permitió al eucariota ancestral la respiración aerobia y con ello aprovechar al máximo la energía contenida en la materia orgánica. Como no es sorprendente en la evolución de un carácter tan antiguo, en varios grupos la mitocondria ha perdido esa capacidad ancestral y a cambio se ha modificado para cumplir otras funciones. También proceden de un evento de endosimbiosis los cloroplastos, en este caso con una cianobacteria, que permiten a las plantas realizar la fotosíntesis. Posteriormente otros grupos de eucariotas consiguieron sus cloroplastos mediante la endosimbiosis secundaria con un alga verde o roja.
Además de la división asexual de las células (mitosis), la mayoría de los eucariontes tiene algún proceso de reproducción sexual basado en la meiosis que no se encuentra entre los procariontes. La reproducción de los eucariontes típicamente implica la existencia de una fase haploide, donde está presente solamente una copia de cada cromosoma en las células, y diploide, donde están presentes dos. Las células diploides surgen por fusión nuclear (fecundación) y las haploides, por meiosis. En los organismos multicelulares, se distinguen tres tipos de ciclos biológicos:
Los organismos unicelulares pueden reproducirse asexualmente por bipartición, gemación o esporulación y sexualmente mediante gametos o por conjugación.[6]
En los eucariontes, la relación de superficie frente a volumen es más pequeña que los procariontes, y así tienen tasas metabólicas más bajas y tiempos de generación más largos.
El origen de la célula eucariota es el proceso biológico más revolucionario desde el origen de la vida desde varios puntos de vista, como es el caso de la morfología, desarrollo evolutivo, estructura genética, relaciones simbióticas y ecología.[7] Todas las células complejas son de este tipo y constituyen la base de casi todos los organismos pluricelulares.[8] Aunque no hay acuerdo sobre cuándo se originaron los eucariotas, en general, se ha sugerido a comienzos del Paleoproterozoico hace unos 2500 millones de años.[9][10] Hasta ahora el fósil más antiguo que puede considerarse eucariota tiene 2200 millones de años y se le conoce como Diskagma el cual representa los primeros indicios de vida pluricelular junto con la biota francevillense de hace 2100 millones de años. La separación entre los eucariotas y su grupo hermano las arqueas Asgard se estimó a finales del Arcaico (periodo Neoarcaico).[10]
Eukarya se relaciona con Archaea desde el punto de vista del ADN nuclear y de la maquinaria genética, y ambos grupos son clasificados a veces juntos en el clado Neomura. Desde otros puntos de vista, tales como por la composición de la membrana, se asemejan más a Bacteria. Se han propuesto para ello tres posibles explicaciones principales:[12][13][14]
Cada vez son mayores las evidencias que parecen demostrar que el origen eucariota es producto de la fusión de una arquea y una bacteria. Mientras el núcleo celular tiene elementos genéticos relacionados con las arqueas, las mitocondrias y la membrana celular tienen características bacterianas. La fusión genética es más evidente al constatar que los genes informativos parecen de origen arqueano y los genes operacionales de origen bacteriano. En todo caso también es cierto que un cierto número de rasgos presentes exclusivamente en los eucariontes son difíciles de explicar por medio de un evento de fusión.[15]
Tampoco está claro el retraso de mil millones de años entre el origen de los eucariotas y su diversificación, pues las bacterias dominaron la biosfera hasta hace unos 800 millones de años.[16] Este intervalo de estabilidad ambiental, litosférica y evolutiva se conoce con el nombre de "boring billion" (aburridos mil millones de años).[17] Este retraso quizá se debiera simplemente a la dificultad de introducción en una biosfera ocupada enteramente por los procariotas. Otras explicaciones están relacionadas con la lenta evolución de los eucariotas o con el aumento del oxígeno, que no alcanzó los niveles ideales para los eucariotas hasta el final de dicho período. El diseño compartimentado de la célula eucariota es el más adecuado para el metabolismo aerobio y es comúnmente aceptado que todos los eucariotas actuales, incluidos los anaerobios, descienden de antecesores aerobios con mitocondrias.[18][19]
Algún tiempo después de que surgiera la primera célula eucariota se produjo una radiación explosiva que las llevó a ocupar la mayoría de los nichos ecológicos disponibles.
La primera célula eucariota era probablemente flagelada, aunque con tendencias ameboides al no tener una cubierta rígida.[21] Desde el antecesor flagelado, algunos grupos perdieron ulteriormente los flagelos, mientras que otros se convirtieron en multiflagelados o ciliados. Cilios y flagelos (incluidos los que tienen los espermatozoides) son estructuras homólogas con nueve dobletes de microtúbulos que se originan a partir de los centriolos.[22]
El carácter ameboide surgió varias veces a lo largo de la evolución de los protistas dando lugar a los diversos tipos de seudópodos de los distintos grupos. El que los ameboides procedan de los flagelados y no al revés, como se pensaba en el pasado, tiene como base estudios moleculares (fusión, partición o duplicación de genes, inserción o borrado de intrones, etc.).[23] [24]
Está generalmente aceptado que los cloroplastos se originaron por endosimbiosis de una cianobacteria y que todas las algas eucariotas evolucionaron en última instancia de antepasados heterótrofos. Se piensa que la diversificación primaria de la célula eucariota tuvo lugar entre los zooflagelados: células predadoras no fotosintéticas con uno o más flagelos para nadar, y a menudo también para generar corrientes de agua con las que capturar a las presas.[21]
En la actualidad hay discrepancia en dónde debe ponerse la raíz del árbol de Eukarya. La posibilidad más aceptada es situarlo entre o próximo a los excavados, que serían el grupo basal de los eucariontes.[3][25]
Durante la primera parte de su historia los eucariontes permanecieron unicelulares. A partir del período Ediacárico los pluricelulares comienzan a profilerar, aunque el proceso con seguridad comenzó bastante antes. Los organismos unicelulares de vida colonial comenzaron a cumplir funciones específicas en una zona del colectivo. Se formaron así los primeros tejidos y órganos. La pluricelularidad se desarrolló independientemente en varios grupos de eucariontes: plantas, hongos, animales, algas rojas, algas pardas y mohos mucilaginosos. A pesar de su pluricelularidad, estos dos últimos grupos se siguen clasificando en el reino Protista.
Las algas verdes, las primeras plantas, se desarrollaron para formar las primeras hojas. En el Silúrico surgen las primeras plantas terrestres y de ellas las plantas vasculares o cormófitas.
Los hongos unicelulares constituyeron filas de células o hifas que agrupadas se convirtieron en organismos pluricelulares absortivos con un marcado micelio. Inicialmente, los hongos fueron acuáticos y probablemente en el período Silúrico apareció el primer hongo terrestre, justo después de la aparición de las primeras plantas terrestres. Estudios moleculares sugieren que los hongos están más relacionados con los animales que con las plantas.
El reino animal comenzó con organismos similares a los actuales poríferos que carecen de verdaderos tejidos. Posteriormente se diversifican para dar lugar a los distintos grupos de invertebrados y vertebrados.
Los eucariontes se dividen tradicionalmente en cuatro reinos: Protista, Plantae, Animalia y Fungi (aunque Cavalier-Smith 2004,[30] 2015[31] reemplaza Protista por dos nuevos reinos, Protozoa y Chromista). Esta clasificación es el punto de vista generalmente aceptado en actualidad, aunque ha de tenerse en cuenta que el reino Protista, definido como los eucariontes que no encajan en ninguno de los otros tres grupos, es parafilético. Por esta razón, la diversidad de los protistas coincide con la diversidad fundamental de los eucariontes.
La reciente clasificación de Adl et al. (2018)[3] evita la clasificación en reinos, sustituyéndola por una acorde con la filogenia actualmente conocida, en la que por otra parte a los clados o taxones no se les atribuye ya categoría alguna, para evitar los inconvenientes que suponen éstas para su posterior actualización. Los principales grupos de esta clasificación (equivalentes a reinos en clasificaciones anteriores) es como sigue:
Algunos grupos de protistas tienen una clasificación dudosa, en particular Cryptista (criptofitas) y Haptista (haptofitas), mientras que otros parecen situarse fuera de los grandes grupos, en particular CRuMs, Ancyromonadida y Hemimastigophora.
Adicionalmente se reconocen dos agrupaciones más grandes. Diaphoretickes (o Corticata) engloba a Archaeplastida y Sar, mientras que Amorphea (o Podiata) agrupa a Amoebozoa y Opisthokonta. Nótese que una forma ameboide o flagelar no indica la pertenencia a un grupo taxonómico concreto, como se creía en clasificaciones tradicionales, creando grupos artificiales desde el punto de visto evolutivo (ver polifilia).
El siguiente árbol filogenético muestras las relaciones entre los principales grupos de Eukarya de acuerdo con Burki et al. 2019:[32]
Eukarya [A] DiaphoretickesHolomycota → ([L] Fungi)
[B] Excavata (P?)Leyendas: [A] Eucarionte heterótrofo y flagelado ancestral. [B] Surco de alimentación ventral. [C] Flagelos heterocontos. [D] Alvéolos corticales. [E] Filopodios. [F] Haptonemas y axopodios. [G] Apéndices en flagelo anterior. [H] Aparición de los cloroplastos por endosimbiosis primaria de una cianobacteria. [I] Fusión triple de genes de la biosíntesis de la pirimidina. [J] Lobopodios. [K] Flagelo posterior. [L] Quitina. [M] Colágeno, blastulación y tejidos diferenciados.
Boenigk2015
En biología y taxonomía, Eukaryota o Eukarya (del griego: εὖ eu —‘bueno’, ‘bien’, 'verdadero'— y κάρυον karyon —‘nuez’, ‘carozo’, ‘núcleo’—) es el dominio (o imperio) que incluye los organismos formados por células con núcleo verdadero. La castellanización adecuada del término es eucariota o eucarionte. Estos organismos constan de una o más células eucariotas, abarcando desde organismos unicelulares hasta verdaderos pluricelulares en los que las diferentes células se especializan para diferentes tareas y que, en general, no pueden sobrevivir de forma aislada.
Pertenecen al dominio o imperio eucariota los reinos de los animales, plantas y hongos, así como varios grupos incluidos en el parafilético reino Protista. Todos ellos presentan semejanzas a nivel molecular (estructura de los lípidos, proteínas y genoma), comparten un origen común, y principalmente, comparten el plan corporal de los eucariotas, muy diferente al de procariotas.
Con excepción de algunos organismos unicelulares, el ciclo de vida eucariota alterna una fase haplonte y otra diplonte, que se consigue mediante la alternancia de meiosis y fecundación, procesos que dan células haplontes y diplontes respectivamente.
Eukarüoodid ehk päristuumsed (Eukaryota) on organismid, kelle rakud on päristuumset (eukarüootset) tüüpi. Eukarüoodid on oma nime saanud selle järgi, et neil asub geneetiline informatsioon (kromosoomidena) rakutuumas, mis on membraaniga ümbritsetud organell.
Eukarüootide kõige olulisemaks tunnuseks ongi membraansete organellide olemasolu rakus, mis võimaldab biokeemilised reaktsioonid ruumiliselt eraldada. Nendeks organellideks on endoplasmaatiline retiikulum ja rakutuum (nende membraanid on omavahel ühenduses), mitokondrid (mõnel eukarüoodil need puuduvad või on taandarenenud), kloroplastid (paljudel puuduvad), Golgi kompleks, lüsosoomid ja peroksüsoomid.
Eukarüootide oluliseks tunnuseks on veel tsütoskelett mikrofilamentide (aktiin) ja mikrotorukeste ehk mikrotuubulite (tubuliin) näol. Aktiinifilamendid määravad raku kuju ja mikrotuubulid tegelevad organellide ümberpaigutamisega, osalevad rakujagunemisel (mitoos ja meioos) ja on viburite koostisosaks. On tõenäoline, et tänapäeva eukarüootide eellasel esines juba tõeline suguline (holomiktiline) sigimine (st. esines meioos). Kui arvestada liigilist mitmekesisust, siis enamik eukarüoote on hulkraksed (koelis-organilise ehitusega).
Eukarüootide päritolu küsimus on veel üsna ebaselge. Kuidas ja millis(t)est prokarüootide rühma(de)st on nad kujunenud? Millal tekkisid esimesed eukarüoodid? Kas eukarüoodid tekkisid umbes samal ajal kui esimesed prokarüoodid (nn. kolm domeeni: Bacteria, Archaea ja Eucarya; Woese et al., 1990), hoopis tunduvalt hiljem ja mingist bakterite (Bacteria sensu Woese) harust (Cavalier-Smith, 2002) või hoopis kahe prokarüoodi ühinemisel (nt. Rivera & Lake, 2004), pole veel päris kindlaks tehtud. Vanimad eukarüootide kivistised on 1,5 (võib-olla isegi 1,7–1,9) miljardit aastat vanad, kuid neid ei ole võimalik kindlalt paigutada ühessegi tänapäeval elavasse rühma (Javaux et al., 2003; Katz et al., 2004).
Eukarüootide evolutsioonis on olulist rolli mänginud endosümbioos. Kahte tüüpi organellid – plastiidid (kloroplastid) ja mitokondrid – pärinevad kunagi eukarüoodi poolt fagotsütoosi teel omandatud bakteritest. Mitokonder pärineb α-proteobakterist ning algselt ilma mitokondrita eukarüoote pole suure tõenäosusega olemas. Kõikidel amitokondriaalsetel (mitokondrita) eukarüootidel on kindlaks tehtud, et neil kas on kunagi olnud mitokonder (tõenduseks on tüüpiliste mitokondri geenide esinemine rakutuumas) või on see tugevalt redutseerunud e. taandarenenud (näiteks hüdrogenosoomideks, mitosoomideks). Kloroplastide evolutsioon on pisut keerulisem. Primaarne endosümbioos (eukarüoot neelab tsüanobakteri, kellest kujuneb sümbiont) leidis aset liitvetikate, punavetikate ja rohevetikate (kellest pärinevad kõik maismaataimed) ühisel eellasel. Sekundaarse endosümbioosi (eukarüoot neelab juba kloroplastiga varustatud eukarüoodi) juhtumeid on olnud rohkem. On kahte tüüpi sekundaarsete plastiididega vetikaid – ühtede plastiidid on pärit punavetikast ja teiste omad rohevetikast. Punavetika tüüpi plastiidid on järgmistel rühmadel: neelvetikad (Cryptophyta), haptofüüdid (Haptophyta), stramenopiilid (Heterokonta; nt. pruunvetikad, ränivetikad) ja vaguviburvetikad (Dinoflagellata). Rohevetika-tüüpi plastiidid on Chlorarachniophyceae (Rhizaria) ja Euglenida (Excavata) esindajate seas. Üldiselt arvatakse, et kaks viimati mainitud vetikarühma omandasid rohevetika sõltumatult. Küsimus, kas punased plastiidid pärinevad ühest eellasest (Chromalveolata hüpotees) või on iga ülal mainitud rühm need omandanud sõltumatult, on veel ebaselge (Katz et al., 2004). Vaguviburvetikate (Dinoflagellata) seas on ette tulnud ka tertsiaarset endosümbioosi (see tähendab sekundaarse plastiidiga eukarüoodi omastamist).
Järgnevalt on toodud eukarüootide suurrühmad ehk "riigid" (Simpson & Roger, 2004), mis hõlmavad enam-vähem kogu eukarüootse mitmekesisuse. Rühma järgi sulgudes on pandud esimeste fossiilide vanused (miljonites aastates, Ma), kes on identifitseeritavad antud rühma liikmena (Javaux et al., 2003).
Taimed, seened ning loomad tekkisid erinevatest ainuraksete eukarüootide rühmadest eelkambriumis (>542 miljonit aastat tagasi).
Hulkraksed loomad ja seened kuuluvad tagaviburiliste (Opisthokonta) hulka. Hulkraksed maismaataimed kuuluvad rühma Plantae (kuhu kuuluvad veel punavetikad, rohevetikad ja liitvetikad). Kõiki eukarüoote, kes ei ole taimed, loomad ega seened, nimetatakse protistideks.
Eukarüoodid ehk päristuumsed (Eukaryota) on organismid, kelle rakud on päristuumset (eukarüootset) tüüpi. Eukarüoodid on oma nime saanud selle järgi, et neil asub geneetiline informatsioon (kromosoomidena) rakutuumas, mis on membraaniga ümbritsetud organell.
Eukarüootide kõige olulisemaks tunnuseks ongi membraansete organellide olemasolu rakus, mis võimaldab biokeemilised reaktsioonid ruumiliselt eraldada. Nendeks organellideks on endoplasmaatiline retiikulum ja rakutuum (nende membraanid on omavahel ühenduses), mitokondrid (mõnel eukarüoodil need puuduvad või on taandarenenud), kloroplastid (paljudel puuduvad), Golgi kompleks, lüsosoomid ja peroksüsoomid.
Eukarüootide oluliseks tunnuseks on veel tsütoskelett mikrofilamentide (aktiin) ja mikrotorukeste ehk mikrotuubulite (tubuliin) näol. Aktiinifilamendid määravad raku kuju ja mikrotuubulid tegelevad organellide ümberpaigutamisega, osalevad rakujagunemisel (mitoos ja meioos) ja on viburite koostisosaks. On tõenäoline, et tänapäeva eukarüootide eellasel esines juba tõeline suguline (holomiktiline) sigimine (st. esines meioos). Kui arvestada liigilist mitmekesisust, siis enamik eukarüoote on hulkraksed (koelis-organilise ehitusega).
Eukariotoak zelula nukleoa eta beste organulu batzuk zelula mintzaz inguratuta duten izaki bizidunak dira. Eukariotoak Eukarya domeinuaren parte dira, eta zelulabakar edo zelulaniztunak izan daitezke. Zelula eukariotoa prokariototik (hau da, bakterioak eta arkeobakterioak) bereizten duen ezaugarria organuluak mintzaz babestuak egotea da, bereziki nukleoa. Nukleo horretan material genetikoa biltzen da, gaineztadura nuklearrren babesean[1][2][3]. Nukleoa izateak ematen die eukariotoei euren izena, antzinako grezierako εὖ (eu, "ondo" edo "benetakoa") eta κάρυον (karyon, "muin", "hazi" edo "mami")[4]. Gaineztadura edo mintzaz babestutako beste organulo batzuk ere badituzte zelula eukariotoek, adibidez mitokrondrioak eta Golgi aparatua. Gainera, landare eta algek kloroplastoak ere badituzte. Arkeobakterio eta bakterioek ez bezala, eukariotoak elkartu eta izaki zelulaniztunak sor ditzakete, ehun eta zelula mota ezberdinekin.
Eukarya domeinuaren barruan Protista, Fungi (onddoak), Plantae (landareak) eta Animalia (animaliak) izeneko erreinuak daude[5][6]. Eukariotoak asexualki ugal daitezke mitosiaren bidez, edo sexualki, meiosi eta gameto fusioaren bidez. Mitosian, zelula zatitzen da eta genetikoki identikoak diren beste bi zelula sortzen ditu. Meiosian, DNAren erreplikazioa bi aldiz egiten du zelula zatiketa, lau zelula haploide sortuz. Zelula hauek sexuaren bidez elkartzen dira, eta gameto izena dute. Gameto bakoitzak kromosoma multzo bakarra du, meiosiaren birkonbinazioan zehar sortutakoak[7].
Eukariotoak monofiletikoak direla ematen du, hau da, jatorri bera duten izakiz osatutako domeinua da. Hiru domeinu sistemako domeinuetako bat da, baina beste biek ez dute goian aipatutako jokamoldea[8]. Eukariotoak izaki bizidun guztien multzoaren zati txiki bat baino ez dira[9].
Zelula eukariotiko gehienak mikroskopikoak badira ere, prokariotikoak baino askoz handiagoak dira: tipikoki hamar aldiz handiagoak luzeraz, eta gutxienez mila aldiz handiagoak bolumenari dagokionez. Badirudi tamaina maximoa elikagaien eta oxigenoaren difusio-tasak mugatzen duela. Tamainarekin batera noski osagai-kopurua ere oso altua da: zelula eukariotiko batean 10.000-20.000 proteina desberdin egon daitezke, eta proteinen kopuru osoa 10.000 milioi inguruko izan daiteke.
Egiturari dagokionez, zelula eukariotiko guztietan bi konpartimentu bereizten dira: zelula nukleoa eta zitoplasma. Zitoplasma gehienetan nukleoa baino handiagoa eta oso ondo antolatuta dago: hemen hainbat organulu eta egitura ezberdin bereizten dira eta prozesu anitz burutzen dira. Organulu zitoplasmatikoak mintz batez mugatutako barrunbeak dira, bakoitzak bere osagai espezifikoekin. Organulu bakoitzean funtzio konkretu bat (edo batzuk) burutzen da: zelula lantegi bat izango balitz organuluak funtzio edo prozesu ezpezifikoetaz arduratzen diren atalak izango lirateke.
Animali zelula batean, organulu zitoplasmatikoek zelularen bolumenaren %50 inguru okupatzen dute. Orokorrean antzeko funtzioak betetzen dituzte zelula mota guztietan baina organulu bakoitzaren kopurua eta zenbait ezaugarri desberdinak dira zelula-motaren arabera. Organuluen garrantziaz ohartzeko kontuan hartu behar dugu organuluen mintzen azalera mintz plasmatikoarena baino 20 aldiz handiagoa izan daitekeela.
Erretikulu endoplasmatikoa (EE) bere izenak adierazten duen moduan sare konplexu bat da hodiz eta sakuluz osatua. Labirinto itxura duen organulu honen mintzak zelularen mintz guztien erdia izan daiteke. Zentro biosintetiko bat da: hemen lipido zelular gehienak eta proteina asko sintetizatu eta modifikatzen dira.
Multzoka agertzen diren zisterna zapalez osatuta dago. Hemen erretikulu endoplasmatikotik datozen produktuen biosintesiak jarraitzen du eta gainera produktu horien sailkapena burutzen da zelularen hemuga desberdinetara bidaltzeko.
Liseri-entzimak gordetzen dituzten poltsak dira. Organulu hauetan molekula eta egitura anitz apurtzen dira; bai endozitosiaren bidez sartzen direnak, bai barneko osagaiak, eliminatuak izan behar direnak.
Endozitaturiko molekulak lisosometara iritsi baino lehen beste barrunbeetatik pasatzen dira, hauek endosomak ditugu: hemen kanpotik datozen produktuak sailkatzen dira.
Mitokondrioak organulu handiak dira, mintz bikoitz batez inguraturik[10]. Hemen arnasketa burutzen da eta ATP molekulak ekoizten dira; gero ATPa zelula osoan erabiliko da hainbat prozesu zelularratan (nukleoan ere bai). Mitokondrioek oso ezaugarri berezia aurkezten dute: bere gai genetiko propioa dauka, proteina mitokondrial batzuk sintetizatzeko ezinbestekoa dena[11].
Entzimak gordetzen dituzten poltsak dira non hainbat erreakzio oxidatibo burutzen diren.
Zitoplasmatik organuluak eliminatzen baditugu geratzen den barrunbeari (eta hor dauden osagaiei) zitosola deitzen diogu. Zitosola askotan zelularen barrunberik handiena da: zelularen bolumenaren erdia baino gehiago hartzen du. Hemen zelularentzat funtsezkoak diren prozesu asko etengabe gertatzen ari dira.
Hasteko esan behar dugu zelularen proteina guztien sintesia zitosolean hasten dela. Hori dela eta erribosomak, itzulpenaz arduratzen diren egiturak zitosolean daude. Animalia zelula batean 10 milioi erribosoma egon daitezke, hauetariko batzuk aske daude eta beste batzuk EE-aren mintzei itsatsita. Gainera proteinen degradazioa ere hemen gertatzen da. Prozesu honetaz arduratzen diren egiturak proteasomak deritze; hauek, erribosomak bezala, elkarte makromolekularrak dira.
Bestalde zitosolean sintesi eta degradazio-erreakzio asko gertatzen ari dira etengabe eta aldiberean. Degradazioa batez ere energia lortzeko (ATP molekuletan metatzen dena) eta sintesia (aminoazido, nukleotido eta gantz azidoen biosintesia), makromolekulen monomeroak sortzeko. Dakigun bezala bide metabolikoak ez dira independenteak, gurutzatzen dira. Erreakzio hauen artean garrantzi berezia dauka glukolisiak: ATP lortzeko bide kataboliko non glukosaren degradazioa gertatzen den. Bestalde zitosola erregaien metalekua da. Zelulak erregaiak behar ditu energia sortzeko. Erregaiak disoluzioan dauden molekula txikiak izan daitezke, glukosa, alegia; baina erregaien erreserbak batez ere egitura ez-solugarrien moduan metatzen dira. Horrelako moduan energia-kopuru handia oso bolumen txikian metatzea lortzen da.
Erreserba-egitura garrantzitsuenak bi dira:
Oso ugariak izan daitezke zenbait zelula-motatan; bereziki lipido-tantek adipozitoetan ia zitoplasma osoa okupatu ahal dute eta organulu bat baino askoz handiagoak izan daitezke. Orokorrean egitura hauek inklusio zitoplasmatikoak deitzen dira: gai espezfikoen metaketak, gehienetan zelularen jarduera biosintetikoaren ondorioz sortuak direnak.
Hasieran ur disoluzioa bat besterik ez zela pentsatu zen; zehazki, ia likidoa den gel moduko disoluzio bat. Disoluzio honetan makromolekulak eta molekula txikiak disolbaturik zeuden. Gaur egun badakigu zitosola oso ondo antolatuta dagoela: sare konplexu bat osatzen duten zuntzez zeharkaturik dago; sare hau zitoeskeletoa da. Zitoeskeletoa beraz, zitosolaren funtsezko osagaia da: zitoeskeletoari esker zitosoleko osagaiak eta barrunbe honetan gertatzen diren erreakzio eta prozesuak espazialki antolatuta daude, hau da, kokapen espezifikoetan agertzen dira. Gainera itxura ematen dio zelulari, mugimendu zelularrez arduratzen da eta funtsezkoa da zatiketa zelularra burutzeko. Zitoeskeletoa firu proteiko ezberdinez osatuta dago; zehazki hiru firu-mota daude:
Zelula eukariotoaren barrunbeak (zelula nukleoa, organulu zitoplasmatikoak eta zitosola) independenteak badira ere, ez dira egitura isolaturik; haien artean ordea elkartrukeak gertatzen ari dira etengabe. Hau ulertzeko kontuan hartu behar dugu barrunbe bakoitzaren osagaiak ez direla sortzen barrunbean bertan, alderantziz, askotan beste lekutan sintetizatzen dira; horregatik mekanismo espezifikoak beharrezkoak dira emandako helmuga batetara heltzeko.
Nukleoaren proteina guztiak esateko zitosolean ekoizten dira eta gero gaineztadura nuklearra zeharkatu behar dute nukleoan sartzeko. Zelulak garraio-sistema desberdinak erabiltzen ditu molekulak barrunbeen artean elkartrukatzeko. Zenbait organulu, erretikulu endoplasmatikoa, Golgiren aparatua, lisosomak eta endosomak hain zuzen, besikulen bidez komunikatuta daude:
Fenomeno honi trafiko besikularra deitzen diogu; oso konplexua da eta oso ondo erregulatuta dago. Garraio-xixkuak beraz, oso ugariak dira edozein animalia zelulatan. Trafiko honetan inplikatutako organuluak etengabe mintz-zatiak elkartrukatzen ari dira, hori dela eta esaten da barne-mintz sistema osatzen dutela[12]. Barnemintz sistematik kanpo mitokondrioak eta peroxisomak geratzen dira, hori dela eta, organulu hauen mintzetan translokadoreak daude osagai espezifikoen inportazioa burutzeko. Bestalde nukleoa eta zitosolaren arteko elkartrukeak nukleoa inguratzen duen gaineztaduraren egituren bidez burutzen da; horiek nukleo-poroak dira.
Zelula eukariotoaren zitoplasma mintz plasmatikoak mugatzen du. Mintz plasmatikoa, egiturari dagokionez, berdina da zelula guztietan, baina eukariotoetan mintz plasmatikoak oso espezializazio-maila altua lortu du. Gogoratu behar dugu prokariotoetan mintz plasmatikoan burutzen diren prozesu asko, ATParen sintesia esateko, eukariotoetan organulu zitoplasmatikoetan burutzen direla. Oro har esan dezakegu eukariotoen mintz plasmatikoa batez ere garraio, komunikazio eta adhesio prozesuetan inplikatuta dagoela, hau da, inguruneariko harremanekin zerikusia duten funtzioetan.
Bestalde animali zeluletan mintz plasmatikoak gaitasun bereziki garrantzitsua garatu da: endozitosi eta exozitosiaren gaitasuna; hau da, gaiak barneratzea edo kanporatzea mintz-zatiak erabiliz. Endozitatzen diren produktuak batzuetan oso handiak dira eta kasu honetan fagozitosiaz hitz egiten dugu. Prozesu hauek burutzeko zelularen azala oso malgua izan behar da, beraz, animalia zelulak biluziak dira, hau da, ez daukate zelula hormarik.
Nukleoan DNA zelular gehiena gordetzen da (baina ez guztia) eta zitoplasmatik banatuta dago gaineztadura nuklearra izeneko bi mintzez osatutako egituraren bidez Egitura honi esker gai genetikoa zitoplasman burutzen den jarduera metabolikotik eta mugimendu zitoplasmatikoetatik babestuta dago[13].
Zelula eukariotikoaren DNA kopurua oso altua da; giza zelula tipiko batek esate baterako, bakterio batek baino 1.000 aldiz DNA gehiago izaten du. Bestalde informazio genetikoa DNA molekula desberdinetan gordetzen da. DNA molekula eukariotikoak oso luzeak dira eta beti proteina espezifikoekin (histonak izeneko proteinekin hain zuzen) elkartuta daude; elkarte honi kromosoma deitzen diogu: DNA molekula bat gehi proteinak. Nukleoan kromosoma batzuk biltzen dira baina zelula zatiketan ez dagoenean hauek ez dira egitura trinkoak eta ez dira bereizten; hori dela eta kromosoma-multzo hau izendatzeko askotan beste termino bat erabiltzen da: kromatina[14][15].
Kromatinaz gain nukleoaren barruan beste egitura bat bereizten da: nukleoloa, erribosomak ekoizteko zentroa. Jarduera nuklearrari dagokionez esan behar dugu nukleoan RNA sintesia burutzen dela baina ez proteinen sintesia; beraz, gene-adierazpenaren bi prozesuak, transkripzioa eta itzulpena, banatuta daude; prokariotoetan ordea aldi berean gertatzen dira.
Ikuspuntu funtzionaletik ere zelula eukariotoa oso konplexua da. Zelula eukariotoaren funtzioak sakonki aztertuko ditugu ondorengo kapituluetan, hemen bakarrik oinarrizko funtzio bat aipatuko dugu: ugalketa. Zelula eukariotoek prozesu bat asmatu dute gai genetikoa banatzeko: mitosia. Mitosian DNA molekulak paketatzen dira egitura trinkoak (kromosoma mitotikoak, bi kromatidez osatuak) sortzeko. Gero, zitoeskeletoari esker, kromosomak bi multzotan banatzen dira bi zelula alaba genetikoki berdinak sortzeko. Mekanismo honen bidez zelula eukariotoak ugaltzen dira. Hau ugalketa asexuala dugu.
Hala ere eukariotoek beste ugalketa-mota asmatu dute non bi zelula batzen dira zelula alaba genetikoki desberdinak sortzeko: ugalketa sexuala meiosiaren bidez. Fenomeno hau ikuspuntu ebolutibotik oso garrantzitsua izan da izaki eukariotikoen aniztasun harrigarria sortzeko[16][17]. Meiosia zelula diploide bakar batetik lau gameto sortzen dituen zelularen zatiketa mekanismoa da. Gametoek parte hartzen duten ugalketa motari deritzo ere, bertako zelula sexualak meiosi bidez lortzen direlarik, ausaz kromosoma homologoek crossing over eginez eta kromosoma ezberdinak lortuz.
Meiosiaren helburua kromosomen erdiak dituzten zelulak sortaraztea da eta bi zatitan bana daiteke prozesua: lehen zatiketa meiotikoa (I. Meiosia) eta bigarren zatiketa meiotikoa (II. Meiosia). Prozesu meiotikoan zelula diploide batetik (2n) bi zatiketen bidez lau zelula haploide (n) sortzen dira. Lehen zatiketan bi zelula diploide lortzen dira (2n) eta bigarrenean bi zelula diploidetik (2n) lau gameto haploideak (n) agertzen dira.
Zelula eukariotoa deskribatzeko animalia zelula eredutzat hartu egin dugu. Landare-zelularen antolamendua funtsean antzekoa da baina zenbait ezaugarri espezifiko aipa daitezke. Tamainari dagokionez gehienetan landare-zelulak animalia zelulak baino handiagoak dira. Bestalde zelularen egiturari dagokionez landare-zelulek osagai espezifiko batzuk izaten dituzte; garrantzitsuenak zelula horma, bakuolak eta kloroplastoak dira. Zelula horma, mintz plasmatikoaren gainetik dagoen egitura zurruna da, bere osagai nagusia zelulosa izanik (glukosaren polimero bat).
Bakuolak likidoz betetako barrunbeak dira, funtzio askotan inplikatuta daudenak. Bat baino gehiago egon daitezke, baina askotan landare-zelularen zitoplasma osoa bakuola handi batez okupatuta dago. Bakuolaren barruan presio osmotikoa oso altua da eta hori dela eta oso garrantzitsua da zelularen bolumena kontrolatzeko. Horrez gain bakuolak erabiltzen dira hainbat molekula metatzeko eta degradazio-prozesuak burutzeko, hau da, bakuolek jarduera lisosomikoa ere aurkezten dute. Kloroplastoak fotosintesia burutzeko erabiltzen diren organulu zitoplasmatikoak dira.
Egiturari dagokionez kloroplastoak eta mitokondrioak oso antzekoak dira: mintz bikoitz batez mugatuta daude, barruan mintz asko daude, tilakoideak, non konplexu fotosintetikoak kokatuta dauden, eta gainera organuluak bere genoma propioa dauka. Konplexu fotosintetikoetan fotosintesia burutzeko entzima eta eragile guztiak bilduta daude, horien artean klorofila pigmentua. Pigmentu hau ezinbestekoa da argi-energia harrapatzeko eta bere presentziagatik kloroplastoak eta hortaz landare-zelulak ere berde kolorekoak dira. Azpimarratu behar da kloroplastoak ez direla agertzen landare-zelula guztietan, bakarrik fotosintesia gertatzen deneko zeluletan; dena den kloroplastoen antzeko organuluak agertzen dira. Izan ere kloroplastoak organulu-familia handi baten partaideak dira; plastoak izeneko familia hain zuzen.
Izaki eukariotiko asko, prokariotoak bezala, zelula bakar batez osatuta daude, hauek eukarioto zelulabakarrak ditugu. Eukarioto zelulabakarrek osatzen duten taldeak protistoak du izena. Protistoak bakterioak baino askoz konplexuagoak dira, hala ere mikroskopikoak dira, beraz, bakterioak bezala, mikroorganismoak dira.
Protistoek aniztasun harrigarria aurkezte dute. Konplexuenak protozooak dira[18]. Protozooek, zelulabakarrak izan arren, animaliak gogorarazten dizkigute: mugikorrak, oso aktiboak, eta asko harrapakariak dira. Nahiko handiak dira (zelula eukariotio tipiko batekin konparatuz) eta euren anatomia oso konplexua da: oso egitura espezializatuak azaltzen dituzte (flageloak, ile sentikorrak, hanka itxurako apendizeak, aho-egiturak, muskuluen antzeko egitura uzkurkorrak, eta abar).
Diatomeak, beste protisto talde bat, ez dira mugikorrak, fotosintetizatzaileak dira, eta zelula silizez osatutako kutxa batean sartuta dago. Legamiak onddoen antzekoak dira (mugiezinak, ezfotosintetizatzaileak) eta egiturari dagokionez oso sinpleak dira; izan ere legamiak zelula eukariotorik sinpleentzat har ditzakegu. Ezaugarri honengatik hain zuzen, legamiak, eta zehazki Saccharomyces cerevisiae legamia (ogia egiteko erabiltzen dena), oso erabilgarriak suertatu dira zelula eukariotoaren prozesu konplexuak ikertzeko: ziklo zelularraren kontrola esate baterako zelula hauetan agerian jartzea lortu da.
Organismo zelulabakarrez gain, zelula eukariotoa izaki multizelularrak sortzeko gai izan da; izaki hauek eboluzioan zehar gero eta konplexuago bihurtu dira goi-mailako animalia eta landareak eman arte. Elkarte zelularrik sinpleenak koloniak dira. Kolonia batean zelula guztiak oso antzekoak dira; gainera kolonia desantola daiteke eta askatzen diren zelulak indibidualki bizi daitezke. Benetako organismo multizelularrak koloniak baino konplexuagoak dira; elkartzen diren zelulek funtzio espezializatuak betetzen baitituzte.
Bestalde organismo zelulanitza elkarte iraunkorra da: zelulak lotuta mantentzen dira haien artean adhesio-mekanismo espezifikoen bidez. Gainera, animalien kasuan, zelulez gai beste elementu bat oso garrantzitsua da: matrize estrazelularra, zelulen artean dagoen gai azelularra. Zelulak eta matrize estrazelularra modu espezifikoetan antolatzen dira ehunak sortzeko. Ehunak funtsean mota bereko zelulez osatuta daude eta matrize estrazelular espezifiko bat aurkezten dute. Oinarrizko ehuna, animalia guztietan agertzen dena, epitelio-ehuna da; izan ere animalia sinpleenak (koralak esate baterako) bakarrik orri epitelialez daude osatuta. Epitelioak orri zelularrak dira non zelulak modu ordenatu batean kokatuta dauden.
Goi-mailako animalietan ehun-mota asko eta zelula-mota asko daude: ugaztunetan 200 zelula-mota desberdin baino gehiago deskribatu egin dira. Zelula-mota bakoitzak bere funtzio espezifikoa izaten du eta hau bere morfologian islatzen da. Argi dago eukariotoek lortu egin duten konplexutasun-maila harrigarria dela. Hala ere ez dugu ahaztu behar, hain izaki konplexuak izan arren, azken finean zelula bakar batetatik sortuak direla, zelula hori zigotoa da. Hau da, organismo osoa garatzeko informazioa zelula eukarioto bakar batean gordetzen da.
Zelula eukariotoak duela mila miloi urte sortu zirela pentsatzen da[19][20]. Lehenengo izaki eukariotoak zelulabakarrak ziren (protistak). Eukariotoen sailkapena asko aldatu da azken urteetan[21][22].
Zelula eukariotikoaren jatorria bizitzaren eboluzioaren barruko mugarri bat izan zen, eukariotoak zelula konplexu eta organismo zelulaniztun guztiak barnebiltzen dituztelako. Hala ere, gertakarien kronologia ezartzea oso ariketa zaila da; Knollek 2006an proposatu zuen orian dela 1.600 eta 2.100 milioi urte artean gertatu zela. Akritarko batzuk orain dela 1.650 milio urte jada existitzen ziren, eta baliteke Grypania alga orain dela 2.100 milioi urte bizi izana[24]. Geosiphonen antza duen Diskagma onddo fosila aurkitu da orain dela 2.200 milioi urteko paleozoru batean[25].
Egitura bizidun antolatuak aurkitu dira Gabongo Francevillian B Formazioko eskisto beltz Paleoproterozoikoan, orain dela 2.100 milioi urtekoak. Bizitza eukariotoa garai horretan eboluzionatzea posible da[26]. Gaur egungo taldeekin harreman argia duten fosilak orain dela 1.200 milioi urte agertu ziren, alga gorrien taldekoak, nahiz eta baliteke Vindhya arroan aurkitu diren filamentu batzuk algak izatea. Hala balitz, orain dela 1.600 edo 1.700 milioi urteko fosilak izango lirateke[27].
Biomarkadoreetan oinarrituta oinarrizko eukariotoak lehenago sortu zirela uste da. Australiako eskistoetan aurkitutako ziklopentanoperhidrofenantrenoa dela eta, orain dela 2.700 milioi urte eukariotoak egon zitezkeela proposatu da[28][29].
Nukleoko DNA eta eukariotoen makinaria genetikoak antza handiagoa du Arkeobakterioekin bakterioekin baino; horregatik, proposamen batzuetan arkeobakterioak Neomura kladoan batzen dira eukariotoekin. Beste eremu batzuetan, hala nola mintzaren osaketa, eukariotoak bakterioen antzekoak dira. Hiru proposamen daude hau azaltzeko:
Beste proposamen alternatiboak ere badaude hau azaltzeko:
Beste inongo talderik ez bada sartzen eukariotoen sorreraren hipotesian, hiru aukera daude bakterio, arkaea eta eukariotoen arteko harremanak emateko, talde monofiletikoak direla pentsatzen badugu. Azpiko taulan batetik hirura arte izendatzen dira. Eozitoaren hipotesia 2. hipotesiaren aldaera bat da, non Archaea parafiletikoa den. Taldeen izenak Harish eta Kurlandek 2017an argitaratutako lanetik hartu dira[40]:
Bizitzaren zuhaitzaren erroarentzat emandako hipotesiak 1 – Bi inperio 2 – Hiru domeinu 3 – Gupta 4 – Eozito AKUArchaea
Bacteria
Eukaryota
Eukaryota
Archaea
Bacteria
Eukaryota
Bacteria
Archaea
Eukaryota
Archaea-Crenarchaeota
Archaea-Euryarchaeota
Bacteria
Eukariotoak zelula nukleoa eta beste organulu batzuk zelula mintzaz inguratuta duten izaki bizidunak dira. Eukariotoak Eukarya domeinuaren parte dira, eta zelulabakar edo zelulaniztunak izan daitezke. Zelula eukariotoa prokariototik (hau da, bakterioak eta arkeobakterioak) bereizten duen ezaugarria organuluak mintzaz babestuak egotea da, bereziki nukleoa. Nukleo horretan material genetikoa biltzen da, gaineztadura nuklearrren babesean. Nukleoa izateak ematen die eukariotoei euren izena, antzinako grezierako εὖ (eu, "ondo" edo "benetakoa") eta κάρυον (karyon, "muin", "hazi" edo "mami"). Gaineztadura edo mintzaz babestutako beste organulo batzuk ere badituzte zelula eukariotoek, adibidez mitokrondrioak eta Golgi aparatua. Gainera, landare eta algek kloroplastoak ere badituzte. Arkeobakterio eta bakterioek ez bezala, eukariotoak elkartu eta izaki zelulaniztunak sor ditzakete, ehun eta zelula mota ezberdinekin.
Eukarya domeinuaren barruan Protista, Fungi (onddoak), Plantae (landareak) eta Animalia (animaliak) izeneko erreinuak daude. Eukariotoak asexualki ugal daitezke mitosiaren bidez, edo sexualki, meiosi eta gameto fusioaren bidez. Mitosian, zelula zatitzen da eta genetikoki identikoak diren beste bi zelula sortzen ditu. Meiosian, DNAren erreplikazioa bi aldiz egiten du zelula zatiketa, lau zelula haploide sortuz. Zelula hauek sexuaren bidez elkartzen dira, eta gameto izena dute. Gameto bakoitzak kromosoma multzo bakarra du, meiosiaren birkonbinazioan zehar sortutakoak.
Eukariotoak monofiletikoak direla ematen du, hau da, jatorri bera duten izakiz osatutako domeinua da. Hiru domeinu sistemako domeinuetako bat da, baina beste biek ez dute goian aipatutako jokamoldea. Eukariotoak izaki bizidun guztien multzoaren zati txiki bat baino ez dira.
Aitotumaiset[1][2] (Eucarya) eli eukaryootit tai aitotumalliset ovat yksi eliöiden kolmesta domeenista. Ne koostuvat solusta tai soluista, joissa on tuma ja monenlaisia erikoistuneita soluelimiä eli soluorganelleja. Ne ovat rakenteeltaan huomattavasti mutkikkaampia ja suurempia kuin esitumalliset bakteerit ja arkeonit.
Koko eliökunta voidaan jakaa esi- ja aitotumaisiin, joista ensimmäiseen kuuluvat bakteerit sekä arkeonit ja jälkimmäiseen kaikki muut. Ongelmana tällaisessa jaossa on kuitenkin se, että arkeonit ja aitotumaiset ovat toisilleen läheisempää sukua kuin bakteereille.
Monisoluiset kasvit, sienet ja eläimet kuuluvat aitotumaisiin. Yksisoluisia eukaryootteja on perinteisesti kutsuttu alkueläimiksi.
Aitotumaisten solujen selvin tunnusmerkki on se, että niillä on selvä tumakalvo, jonka sisällä solun perintöaines on. Tumakalvon vuoksi aitotumaiset solut kuljettavat lähetti-RNA:n eli mRNA:n ulos tumasta ribosomeille, jotta nämä voisivat aloittaa solulimassa tapahtuvan proteiinisynteesin.
Aitotumaisilla on tuman lisäksi muita erilaistuneita, kalvon ympäröimiä soluelimiä. Niiden uintisiimat ovat paljon monimutkaisempia kuin esitumallisilla tavattavat. Aitotumaisilla nämä siimat muodostuvat tosiinsa limittyvistä mikrotubuluksista.
Bakteerit, arkit ja aitotumaiset ovat ilmeisesti polveutuneet samasta kantamuodosta. Nämä ryhmät ovat kuitenkin eronneet toisistaan erittäin varhaisessa vaiheessa. Ensin bakteerit eriytyivät omaksi kehityslinjakseen. Arkit ja aitotumaiset erosivat toisistaan myöhemmin.
Aitotumaisten syntyä on väitetty jopa elämän syntyä merkittävämmäksi harppaukseksi elämän historiassa. Aitotumaisia väitetään jopa 2,7 miljardia vuotta vanhoiksi, mutta väite on kiistanalainen ja perustuu kemiallisiin jäämiin eli ns. biomarkkereihin ja kivimuodostelmiin, akritarkkeihin, joita on vaikeaa tulkita. Ensimmäiset nykyisenkaltaisiin aitotumaisiin viittaavat fossiilit on tavattu 1,2 miljardin vuoden takaa. Aitotumaisia oli olemassa kuitenkin viimeistään 800 miljoonaa vuotta sitten.
Aitotumaisten eliöiden tuma ja kromosomisto kehittyivät, kun eliöiden genomit olivat kasvaneet liian suuriksi, jotta yksittäisen rengasmaiseen kromosomiin perustuva rakenne olisi toiminut tehokkaasti. Kromosomisto puolestaan edellytti mitoosin kehittymistä.
Tumakalvo, uintisiimat ja mitokondriot ovat ilmeisesti kehittyneet jo ennen kromosomiston syntymistä. Aitotumaisen solun uskotaan syntyneen, kun aitotumaisten kantamuoto otti soluihinsa endosymbiontiksi ilmeisesti purppurabakteerien kehityslinjaan kuuluneet esitumallisen mikrobin. Tällä tavoin isäntäsolu sai kyvyn soluhengitykseen. Mitokondriot ovat tämän käsityksen mukaan näitä taantuneita bakteereja. Kasvien ja viherlevien viherhiukkaset ovat vastaavasti taantuneita endosymbionttisia sinileviä. On todettu, että nykyäänkin on yksisoluisia, jotka elävät bakteerien ja sinilevien tai toisten yhteyttävien aitotumaisten kanssa symbioosissa.
soluelimet · solukalvo · solulima · solun tukiranka · soluseinä · tuma
Soluelimetdiktyosomi · endoplasmakalvosto · Golgin laite · lysosomi · mikrovillus · mitokondrio · peroksisomi · proteasomi · ribosomi · vakuoli · vesikkeli · viherhiukkanen
Solun tukirankamikrofilamentit · mikrotubulukset · välikokoiset säikeet
Tuma Muita osiaAitotumaiset (Eucarya) eli eukaryootit tai aitotumalliset ovat yksi eliöiden kolmesta domeenista. Ne koostuvat solusta tai soluista, joissa on tuma ja monenlaisia erikoistuneita soluelimiä eli soluorganelleja. Ne ovat rakenteeltaan huomattavasti mutkikkaampia ja suurempia kuin esitumalliset bakteerit ja arkeonit.
Koko eliökunta voidaan jakaa esi- ja aitotumaisiin, joista ensimmäiseen kuuluvat bakteerit sekä arkeonit ja jälkimmäiseen kaikki muut. Ongelmana tällaisessa jaossa on kuitenkin se, että arkeonit ja aitotumaiset ovat toisilleen läheisempää sukua kuin bakteereille.
Monisoluiset kasvit, sienet ja eläimet kuuluvat aitotumaisiin. Yksisoluisia eukaryootteja on perinteisesti kutsuttu alkueläimiksi.
Eucaryotes
Les eucaryotes (Eukaryota) sont un domaine regroupant tous les organismes, unicellulaires ou multicellulaires, qui se caractérisent par la présence d'un noyau et généralement d'organites spécialisés dans la respiration, en particulier mitochondries chez les aérobies mais aussi hydrogénosomes chez certains anaérobies. On le distingue classiquement des deux autres domaines que sont les bactéries et les archées (mais le clade des eucaryotes s'embranche en fait parmi ces Archées).
Les eucaryotes rassemblent trois grands règnes du monde du vivant : les animaux, les champignons, les plantes, et d'autres (par exemple les algues brunes). Les eucaryotes unicellulaires sont parfois regroupés sous le terme de « protistes » et les non-Eukaryotes sous la dénomination de « procaryotes » (ces deux derniers groupes étant paraphylétiques).
Les eucaryotes peuvent se reproduire de manière sexuée (par méiose et fusion de gamètes) ou non (par mitose). Dans la mitose, une cellule se divise pour produire deux cellules génétiquement identiques. Dans la méiose, la réplication de l'ADN est suivie de deux cycles de division cellulaire pour produire quatre cellules filles haploïdes. Ceux-ci agissent comme des cellules sexuelles (gamètes). Chaque gamète ne possède qu'un seul ensemble de chromosomes, chacun étant un mélange unique de la paire correspondante de chromosomes parentaux résultant d'une recombinaison génétique au cours de la méiose.
Le terme Eukaryota[1],[2],[3] provient du grec eu, « bien » et karuon, « noyau ». Il signifie donc littéralement « ceux qui possèdent un véritable noyau ». Il s'oppose au concept de Prokaryota.
Les eucaryotes forment traditionnellement un empire du monde vivant, ou un domaine dans la classification proposée par Carl Woese. À cette occasion, ce dernier suggère un changement de nom pour Eucarya[4], un terme aujourd'hui très peu employé, en dehors de quelques microbiologistes[5].
Le terme est aussi écrit sous la variante Eukarya[6], notamment par certains biologistes qui, à l'instar de Margulis et Chapman (2009)[7], considèrent le taxon comme un super-règne.
Les cellules eucaryotes possèdent, par opposition aux procaryotes (archées et bactéries) :
L'espèce humaine.
Une paramécie.
Une fougère.
Un champignon : l'amanite tue-mouche.
Les plus anciens eucaryotes attestés seraient âgés de 1,6 Ga, certains acritarches dateraient approximativement de cette époque. Leur origine, toutefois, pourrait être encore plus ancienne. Grypania, vieille de 2,1 Ga, a été rapprochée des algues[9], et les Gabonionta, dans les formations de schistes noirs du Gabon, aussi anciens, suggèrent qu'une vie organisée faisant penser aux eucaryotes existait déjà[10]. L'apparition des eucaryotes est encore plus ancienne. La présence de stérane, marqueur biochimique des eucaryotes dans des formations schisteuses australiennes suggèrent qu'à l'époque deux lignées s'étaient déjà différenciées il y a 2,7 Ga[11].
Les groupes modernes ont d'abord été retrouvés dans les archives fossiles il y a 1,2 Ga sous la forme d'une algue rouge. Mais là aussi, les origines sont plus anciennes puisqu'un fossile trouvé dans le bassin du Vindhya en Inde et datant de 1,6 Ga pourrait bien être une algue filamenteuse[12]. D'autres cellules fossilisées datées de 1,6 milliard d'années et présentant des cellules compartimentées et des organelles ont été découvertes dans des roches sédimentaires en Inde centrale[13]. Il semble y avoir deux types d'algues rouges nommées Rafatazmia chitrakootensis (filamenteuse et contenant de grands disques rhomboïdaux qui pourraient être des restes de chloroplastes) et Ramathallus lobatus (plus globulaire et charnue). Mieux dater l'apparition des premiers eucaryotes est important pour évaluer les vitesse et taux de mutations du génome dans le temps. Faute d'ADN, les chercheurs ne peuvent pas certifier qu'il s'agit d'algues rouges[14].
Dans le monde des bactéries et des archées, le groupe le plus proche des eucaryotes est un super-embranchement d'archées, les archées d'Asgård[15]. Leur génome code une série de protéines identiques ou similaires à des protéines qu'on pensait spécifiques des eucaryotes, et notamment l'actine qui forme le cytosquelette[16]. Au sein des Asgards, l'embranchement le plus proche des eucaryotes est celui des Heimdallarchaeota[17].
La mitochondrie serait le résultat de l'endosymbiose d'une alpha-protéobactérie (une rhodobactérie) par une cellule eucaryote primitive[18].
L'existence de gènes d'endosymbiotes (transférés au noyau de la cellule hôte et intégrés dans le génome de cette dernière) ou de leurs vestiges (demeurant dans le noyau alors que les organites eux-mêmes sont perdus ou dégénérés) révèle que les ancêtres d'eucaryotes dépourvus de mitochondries ont contenu jadis de tels organites[19].
Les eucaryotes comprennent deux clades[20] :
Cladogramme selon les études de Cavalier-Smith, Brown Heiss et Torruella:[21],[22],[23],[24]
Eukaryota DiphodaEucaryotes
Les eucaryotes (Eukaryota) sont un domaine regroupant tous les organismes, unicellulaires ou multicellulaires, qui se caractérisent par la présence d'un noyau et généralement d'organites spécialisés dans la respiration, en particulier mitochondries chez les aérobies mais aussi hydrogénosomes chez certains anaérobies. On le distingue classiquement des deux autres domaines que sont les bactéries et les archées (mais le clade des eucaryotes s'embranche en fait parmi ces Archées).
Les eucaryotes rassemblent trois grands règnes du monde du vivant : les animaux, les champignons, les plantes, et d'autres (par exemple les algues brunes). Les eucaryotes unicellulaires sont parfois regroupés sous le terme de « protistes » et les non-Eukaryotes sous la dénomination de « procaryotes » (ces deux derniers groupes étant paraphylétiques).
Les eucaryotes peuvent se reproduire de manière sexuée (par méiose et fusion de gamètes) ou non (par mitose). Dans la mitose, une cellule se divise pour produire deux cellules génétiquement identiques. Dans la méiose, la réplication de l'ADN est suivie de deux cycles de division cellulaire pour produire quatre cellules filles haploïdes. Ceux-ci agissent comme des cellules sexuelles (gamètes). Chaque gamète ne possède qu'un seul ensemble de chromosomes, chacun étant un mélange unique de la paire correspondante de chromosomes parentaux résultant d'une recombinaison génétique au cours de la méiose.
Tugtar eocaróit ar ainmhithe, plandaí, fungais agus prótaistigh, .i. na beothaigh go léir ina n-eagraítear a gcuid ceall i struchtúir choimpléascacha iniata i scannáin.
An núicléas an struchtúr scannáncheangailte a léiríonn an difríocht idir chealla eocarótacha agus cealla prócarótacha; is mar gheall ar láithreacht an núicléis a húsáidtear an t-ainm "eocarót", ó na focail Gréigise ευ (a chiallaionn "maith/fíor") agus κάρυον ("cnó"). Bíonn orgánaidí scannáncheangailte eile ann i mórán ceall eocarótach, ar nós miteacoindrí, coimpléasc Golgi agus coirpiní Golgi. Is minic a bhíonn lascnaidí uathúla ag eocaróit, déanta de micrifhidíní i leagan amach 9+2.
Os eucariontes son os organismos con células que conteñen un núcleo e outros orgánulos rodeados de membranas. Os eucariontes pertencen ao dominio Eukarya ou Eukaryota. A característica que os distingue dos procariotas (as bacterias e as arqueas) é a posibilidade de que teñan orgánulos rodeados de membrana, especialmente un núcleo celular e que contén o material xenético rodeado dunha membrana nuclear.[1][2][3] Estes organismos constan dunha ou máis células eucariotas, abranguendo dende organismos unicelulares ata verdadeiros pluricelulares nos cales as diferentes células especialízanse en diferentes tarefas e que, en xeral, non poden sobrevivir de forma illada.
Pertencen ó dominio Eukarya os animais, as plantas, os fungos, así como varios grupos denominados colectivamente protistas. Todos eles presentan semellanzas a nivel molecular (estrutura dos lípidos, proteínas e xenoma) e comparten unha orixe común. Os eucariontes orixináronse hai 2,7 mil millóns de anos, despois de entre 1-1,5 mil millóns de anos de evolución dos procariotas.[4] Os fósiles máis temperáns, como os acritarco son difíciles de interpretar. Formas que poden relacionarse inequivocamente cos grupos modernos comezaron a xurdir hai uns 800 millóns de anos, mentres que a maioría dos grupos fósiles xa se coñecen dende o final do Cámbrico, hai uns 500 millóns de anos.
Os eucariontes repártense entre catro reinos (Protista, Plantae, Animalia e Fungi), cinco, segundo Cavalier-Smith (os anteriores máis o reino Chromista), ou moitos máis (algúns consideran, pero non é punto de vista xeralmente aceptado, que é máis axeitado clasificalos en ata vinte reinos).
O reino Protista é un grupo parafilético que xunta aos eucariontes que non encaixan en ningún dos tres grupos; por esta razón a súa diversidade coincide coa diversidade fundamental dos eucariontes. A recente clasificación de Adl et al. (2005) evita a clasificación en reinos, substituíndoa por unha acorde coa filoxenia actualmente coñecida, na que por outra parte aos taxons non se lles atribúe xa categoría algunha, para evitar os inconvenientes que supón estas para a súa posterior actualización. Esta clasificación sería a seguinte:
Pódese ver que unha forma ameboide ou flaxelar non indica a pertenza a un grupo taxonómico concreto, como se pensaba en clasificacións tradicionais, creando grupos artificiais dende o punto de vista evolutivo (ver parafilia).
Os eucariontes son os organismos con células que conteñen un núcleo e outros orgánulos rodeados de membranas. Os eucariontes pertencen ao dominio Eukarya ou Eukaryota. A característica que os distingue dos procariotas (as bacterias e as arqueas) é a posibilidade de que teñan orgánulos rodeados de membrana, especialmente un núcleo celular e que contén o material xenético rodeado dunha membrana nuclear. Estes organismos constan dunha ou máis células eucariotas, abranguendo dende organismos unicelulares ata verdadeiros pluricelulares nos cales as diferentes células especialízanse en diferentes tarefas e que, en xeral, non poden sobrevivir de forma illada.
Eukarioti su organizmi odnosno stanice kod kojih je nasljedni materijal smješten u jezgri obavijenoj posebnom dvostrukom jezgrinom membranom. U eukariotskoj stanici razvili su se i brojni stanični organeli kojih nema kod prokariotskih organizama, među kojima su: endoplazmatiski retikulum, Golgijev aparat (ili Golgijevo tijelo), lizosomi i dr.
Životinje, biljke, gljive i protisti su eukarioti, dok su bakterije i cijanobakterije prokarioti. Izuzev protista, svi drugi eukarioti su višestanični. Za razliku od eukariota, prokarioti nemaju jezgru i ostale složene stanične strukture. Izraz „eukariot“ potječe od grčke riječi ευ, koja znači pravi/dobar, te od κάρυον, koja znači lješnjak ili jezgru.
Eukariotske stanice su većinom puno veće od onih od prokariota. One imaju niz unutrašnjih membrana i struktura, zvanih organeli, i citoskeleton sastavljen od mikrotubula koji igraju važnu ulogu u definiranju organizacije i oblika stanice. Eukariotska DNK je podijeljena u nekoliko kromosoma, koji su odvojeni mikrotubularnom vretenom tijekom razdiobe nukleusa. Uz dodatak aseksualne stanične diobe, mitoze, većina eukariota ima nekakav proces seksualnog razmnožavanja uz pomoć fuzije stanica, mejoze, koji se ne nalazi u prokariota.
Eukariotske stanice uključuju niz membranom omeđenih struktura, koje se nazivaju endomembranski sustav. Jednostavni odjeljci, koji se zovu vakuole ili vesikule, mogu se formirati pupanjem drugih membrana. Mnoge stanice probavljaju hranu i druge materijale uz pomoć procesa zvanog endocitoza.
Jezgra je okružena dvostrukom membranom sa porama koje dopuštaju materijal da se giba unutra i van. Razna cijevkasta produženja središnje membrane stvaraju ono što se zove endoplazmatski retikulum ili ER, koji je uključen u prenošenje i dozrijevanje proteina. To uključuje grubi ER gdje se ribosomi zakvače, a proteini koji se sintetiziraju ulaze u unutrašnji prostor. Oni većinom ulaze u vesikule, koje se stvaraju od glatkog ER. U većini eukariota, ova vesikula koja prenosi proteine biva modifircirana u nakupinu izravnanih vesikula, zvanih Golgijev aparat.
Vesikule se specijaliziraju za razne upotrebe. Primjerice, lizosomi sadrže enzime koji razgrađuju određeni sadržaj vakuola hrane, a peroksisomi se iskorištavaju da razgrade peroksid koji je inače otrovan. Mnogi protociti imaju kontraktilne vakuole koje skupljaju i izbacuju prekomjernu vodu.
Mitohondriji su organele koje se nalaze u gotovo svim eukariotima. Obavijeni su dvostrukom membranom, od koje se unutrašnja savija u cristae, gdje se odvija aerobno disanje. Oni sadrže svoju vlastitu DNK te se formiraju fizijom od drugih mitohondrija. Smatra se da su se razvili od endosimbiotičnih prokariota. Malen broj protocita koji nemaju mitohondrije sadrže organele, kao što su hidrogenosomi mitozomi.
Biljke imaju i plastide, koji pak imaju svoju vlastitu DNK. Oni obično zauzimaju oblik kloroplasta, koji poput cijanobakterija sadrže klorofil i stvaraju energiju putem fotosinteze. Druge se uključene u spremanje hrane.
Mnogi eukarioti imaju Flagellum. Oni su sastavljeni većinom od tubulina i silija, koji su uključeni u pokret, hranjenje i osjet. Oni se potpuno razlikuju od prokariotskih flagellum. Njih podupire skupina mikrotubula koji izlaze iz centriola.
Centriole su prisutne čak i u stanicama i nakupinama koje nemaju flagellum. Oni se većinom javljaju u skupni od jedno ili dvoje, zvanih kinetidi. Oni formiraju primarnu komponentu u strukturi citoskeletona, te se većinom sastavljaju tijekom više staničnih dioba, gdje se jedna flagela dobiva od roditelja a drugi se deriviraju od njih. Važnost strukture citoskeletona leži u utvrđivanju oblika stanica.
Dioba jezgre se često koordinira sa diobom stanica, ovo se većinom odvija u mitozi, proces koji dopušta svakom novom jezgrom da dobije jednu kopiju od svakog kromosoma. U većini eukariota tu je istodobno i proces seksualnog razmnožavanja, koji tipično uključuje izmjenjivanje između haploidnih generacija, gdje je samo jedna kopija od svakog kromosoma prisutna, i diploidna generacija, gdje su dvije prisutne, koji se odvijaju kroz stapanje jezgre i mejoze. Ipak, postoji i značajna varijacija ovog modela.
Pojava eukariota je bio prijelomni trenutak u evoluciji života, pošto one uključuju sve složene stanice i višestanične organizme. Teško je utvrditi pojavu tog događaja; Knoll sugerira da su se razvili prije oko 1.6 do 2.1 milijarde godina. Fosili koji su povezani uz tu grupu počeli su se javljati prije oko 1.2 milijarde godina, u obliku crvenih algi.
Porijeklo endomembranskog sustava i mitohondrija je također sporno. Fagotrofična hipoteza tvrdi da su membrane nastale razvojem endocitoze te su se kasnije specijalizirale; mitohondriji su dobiveni gutanjem, kao plastidi. Sintrofična hipoteza pak tvrdi da su se proto-eukarioti oslonili na proto-mitohondrije za hranu, te su ih tako okružili; membrane su se razvile kasnije, zahvaljujući mitohondrijskim genima.
Actinopoda, Alveolata, Ambonosphaera, Amoebozoa, Animalia (Linnaeus 1758), Apusozoa, Archaeplastida, Biliphyta Cavalier-Smith 1981, Centroheliozoa, Chlorobiota, Chromalveolata, Chrysophyta, Cryptophyta, Ctenidodinium, Dapcodinium, Dichadogonyaulax, Euglenozoa, Excavata, Fornicata, Fungi, Glaucocystophyceae, Haptophyceae, Heterokontophyta, Heterolobosea, Jakobida, Katablepharidophyta, Malawimonadidae, Meiourogonyaulax, Mendicodinium, Opisthokonta, Oxymonadida, Parabasalia, Paragonyaulacysta, Pareodinia, Plantae, Protozoa, Rhizaria, Rhizopodea, Rhodophyta, Stramenopila, Subtilisphaera, Viridiplantae, neklasificirani eukaryoti
Eukarioti su organizmi odnosno stanice kod kojih je nasljedni materijal smješten u jezgri obavijenoj posebnom dvostrukom jezgrinom membranom. U eukariotskoj stanici razvili su se i brojni stanični organeli kojih nema kod prokariotskih organizama, među kojima su: endoplazmatiski retikulum, Golgijev aparat (ili Golgijevo tijelo), lizosomi i dr.
Životinje, biljke, gljive i protisti su eukarioti, dok su bakterije i cijanobakterije prokarioti. Izuzev protista, svi drugi eukarioti su višestanični. Za razliku od eukariota, prokarioti nemaju jezgru i ostale složene stanične strukture. Izraz „eukariot“ potječe od grčke riječi ευ, koja znači pravi/dobar, te od κάρυον, koja znači lješnjak ili jezgru.
Eukaryoty (Eucaryota; wot starogrjekskeho εὖ eu „dobry“, „woprawdźity“ a κάρυον karyon „worjech“, „jadro“) tworja jednu z třoch domenow w biologiji. Su žiwochi z woprawdźitym bańkowym jadrom, kotrež wobsahuja genetisku maćiznu we formje desoksyribo-nukleinoweje kisaliny (skrót.: DNA, němsce DNS).
Eukaryoty su zwjetša wo wjelekróćne wjetše hač prokaryoty (něhdźe 10-100 raz). Zo by hładke fungowanje celularnych wotběhow nad wjetšimi dalokosćemi nutřka bańki zmóžniło, je wyši organizaciska měra a rozdźělenje ruma bańki do kompartimentow (dźělne wotrězki) ale tež transport mjezy tutymi kompartimentami nuzne. Tuteje přičiny dla su eukaryotiske bańki z pomocu bańkowych organelow strukturowane, kotrež kaž organy ćěła rozdźělne funkcije spjelnja. Najbóle znata organela je bańkowe jadro, z hłownym podźělom genetiskeje maćizny žiwocha. Dalše geny wustupuja po kóždej družinje w mitochondrijach a plastidach.
Dalša wosebitosć eukaryotow leži w proteinowej biosyntezy: Hinak hač prokaryoty su eukaryoty kmane, z samsneje DNA-informacije přez alternatiwny splicing rozdźělne proteiny zhotowjeć.
Struktura a forma so eukaryotiskej bańce přez cytoskelet da. Bu mjez druhim z mikrotubulusow, aktinowych włokninow a mikrofilamentow natwarjeny.
Eukaryoty bu tradicionelnje do swětow wjacebańkowych zwěrjatow, rostlin a hribow ale tež jednobańkowych abo wjacebańkowych protistow rozdźělene. Dokelž protisty pak žane monofyletisku skupinu njetworja a druhe swěty poprawom z rozdźělnych protistowych skupin nastajachu, njeje rozdźělenje do swětow wjace dźeržomna.
Tón čas aktuelna Systematika eukaryotow bu wot Adl et al. 2005 wudźěłana.[1] Woni rozrjaduja eukaryotow do šěsć skupin:
Jeli sy jedyn z mjenowanych njedostatkow skorigował(a), wotstroń prošu potrjecheny parameter předłohi {{Předźěłuj}}. Podrobnosće namakaš w dokumentaciji.
Eukaryoty (Eucaryota; wot starogrjekskeho εὖ eu „dobry“, „woprawdźity“ a κάρυον karyon „worjech“, „jadro“) tworja jednu z třoch domenow w biologiji. Su žiwochi z woprawdźitym bańkowym jadrom, kotrež wobsahuja genetisku maćiznu we formje desoksyribo-nukleinoweje kisaliny (skrót.: DNA, němsce DNS).
Eukaryoty su zwjetša wo wjelekróćne wjetše hač prokaryoty (něhdźe 10-100 raz). Zo by hładke fungowanje celularnych wotběhow nad wjetšimi dalokosćemi nutřka bańki zmóžniło, je wyši organizaciska měra a rozdźělenje ruma bańki do kompartimentow (dźělne wotrězki) ale tež transport mjezy tutymi kompartimentami nuzne. Tuteje přičiny dla su eukaryotiske bańki z pomocu bańkowych organelow strukturowane, kotrež kaž organy ćěła rozdźělne funkcije spjelnja. Najbóle znata organela je bańkowe jadro, z hłownym podźělom genetiskeje maćizny žiwocha. Dalše geny wustupuja po kóždej družinje w mitochondrijach a plastidach.
Dalša wosebitosć eukaryotow leži w proteinowej biosyntezy: Hinak hač prokaryoty su eukaryoty kmane, z samsneje DNA-informacije přez alternatiwny splicing rozdźělne proteiny zhotowjeć.
Struktura a forma so eukaryotiskej bańce přez cytoskelet da. Bu mjez druhim z mikrotubulusow, aktinowych włokninow a mikrofilamentow natwarjeny.
Eukariota adalah organisme dengan sel yang memiliki nukleus dan organel bermembran lainnya.
Eukariota berada dalam takson Eukarya atau Eukaryota. Hal yang membuat sel eukariotik berbeda dari sel prokariotik (Bacteria dan Archaea) adalah bahwa mereka memiliki organel bermembran, terutama inti, yang berisi materi genetik, dan terbungkus oleh selubung nukleus.[3][4][5] Nama Eukaryota digunakan karena adanya inti, yang berasal dari bahasa Yunani ευ (eu, "baik") dan κάρυον (karyon, "kacang" atau "kernel").[6] Sel eukariotik juga mengandung organel bermembran lainnya seperti mitokondria dan badan Golgi. Selain itu, tumbuhan dan alga mengandung kloroplas. Organisme eukariotik bisa berbentuk uniseluler atau multiseluler. Hanya eukariota yang memiliki banyak jenis jaringan yang terdiri dari jenis sel yang berbeda.
Eukariota dapat bereproduksi baik melalui reproduksi aseksual melalui mitosis dan reproduksi seksual melalui meiosis. Dalam mitosis, satu sel membelah untuk menghasilkan dua sel yang identik secara genetik. Dalam meiosis, replikasi DNA diikuti oleh dua putaran pembelahan sel untuk menghasilkan empat sel anak masing-masing dengan setengah jumlah kromosom dari sel induknya (sel haploid). Sel-sel ini bertindak sebagai sel kelamin (gamet – masing-masing gamet hanya memiliki satu pelengkap kromosom, masing-masing campuran unik dari pasangan kromosom orang tua yang sesuai) yang dihasilkan dari rekombinasi genetik selama meiosis.
Domain Eukaryota tampaknya monofiletik, dan membuat salah satu dari tiga domain kehidupan. Dua domain lainnya, Bacteria dan Archaea, adalah prokariota[7] dan tidak memiliki ciri-ciri di atas. Eukariota merupakan minoritas kecil dari semua makhluk hidup.[8] Namun, karena ukuran mereka jauh lebih besar, biomassa kolektif eukariota di seluruh dunia diperkirakan sekitar setara dengan prokariota.[8] Eukariota pertama kali berkembang sekitar 1,6-2,1 miliar tahun yang lalu.
Pada umumnya, sel eukariota memiliki ukuran yang lebih besar dari prokariota dan memiliki bagian-bagian sub-seluler yang disebut dengan organel dan sitoskeleton yang terdiri atas mikrotubulus, mikrofilamen dan filamen antara. Berbeda dengan prokariota, DNA eukariota disimpan dalam kumpulan kromosom yang tersimpan di dalam nuklei yang terbungkus membran nuklei. Selain melakukan pembelahan sel secara aseksual, kebanyakan eukariota juga bisa melakukan reproduksi seksual melalui proses fusi sel, yang tidak ditemukan pada prokariota.
Sel eukariotik memiliki bermacam-macam struktur yang dibatasi membran, yang secara kolektif disebut sistem endomembran. Ruang sederhana, yang disebut vesikel atau vakuola, dapat terbentuk dengan pemisahan dari membran lain. Banyak sel menelan makanan dan bahan lain melalui proses yang disebut endositosis, di mana membran luar melekuk ke dalam kemudian putus membentuk vesikel. Kemungkinan banyak organel bermembran lainnya berasal dari vesikel yang demikian.
Inti sel dilapisi oleh membran ganda, yang memiliki pori-pori yang memungkinkan bahan-bahan keluar-masuk. Bermacam peluasan membran nukleus yang berbentuk tabung atau lembaran membentuk retikulum endoplasma (atau RE), yang terlibat dalam transpor dan pematangan protein. RE terdiri atas RE kasar yang memiliki ribosom yang melekat, dan protein yang disintesis ribosom itu memasuki ruang dalam atau lumen . Kemudian, biasanya mereka memasuki vesikel, yang terpisah dari RE halus. Pada kebanyakan eukariota, vesikel pembawa protein ini dilepaskan dan dimodifikasi pada tumpukan vesikel yang memipih yang disebut badan Golgi atau diktiosom.
Vesikel dapat berspesialisasi untuk beragam kegunaan. Contohnya, lisosom mempunyai enzim yang menguraikan isi vakuola makanan, dan peroksisom yang fungsinya menguraikan peroksida, yang beracun. banyak protozoa memiliki vakuola kontraktil, yang mengumpulkan dan membuang kelebihan air dan ekstrusom, yang mengeluarkan bahan yang dipakai untuk melawan pemangsa atau menangkap mangsa. Pada tumbuhan tingkat tinggi, sebagian besar ruang sel diisi oleh vakuola pusat, yang fungsi utamanya untuk menjaga tekanan osmotik.
Mitokondria adalah organel yang ditemukan pada hampir semua eukariota. Mitokondria diselubungi membran ganda, yang membran dalamnya berlekuk-lekuk ke dalam membentuk krista, tempat berlangsungnya respirasi aerobik. Mitokondria memiliki DNA dan ribosom-nya sendiri dan hanya terbentuk dari pembelahan mitokondria lain. Sekarang mereka umumnya berkembang dari prokariota yang berendosimbiosis, mungkin proteobacteria. Beberapa protozoa yang tidak memiliki mitokondria ditemukan mempunyai organel yang diturunkan dari mitokondira seperti hidrogenosom dan mitosom.
Tumbuhan dan berbagai kelompok alga juga memiliki plastida. Dan plastida ini juga mempunyai DNA sendiri dan berkembang dari proses endosimbiosis, dalam hal ini cyanobacteria. Biasanya plastida berbentuk kloroplas, yang mengandung klorofil dan menghasilkan energi melalui fotosintesis seperti halnya cyanobacteria. Plastida lain terlibat dalam menyimpan makanan. Meskipun plastida mungkin memiliki satu asal, tidak semua grup yang memiliki plastida berkerabat dekat. beberapa eukariota mendapatkannya dari yang lain dengan endosimbiosis penelanan sekunder.
Sel eukariotik mempunyai dua buah isoenzim malate dehydrogenase yang berupa mitokondria (m-MDH) dan plastida (bahasa Inggris: cytoplasmic) (s-MDH). Enzim ini berfungsi untuk konfigurasi isomer L pada asam malik (bahasa Inggris: malate). Inhibitor enzim ini adalah ATP, ADP, AMP, tiroksin, yodium sianida, and molekul yodium.[9]
Banyak eukariota memiliki tonjolan sitoplasma motil yang panjang, yang disebut flagela, atau struktur yang mirip yang disebut silia. Flagela dan silia kadang-kadang disebut sebagai undulipodia,[10] dan terlibat dalam pergerakan, makan, dan sensasi. Flagela dan silia tersusun terutama dari tubulin. Mereka sangat berbeda dari flagela prokariotik. Mereka didukung oleh mikrotubulus yang muncul dari badan basal, disebut juga kinetosom atau sentriol, secara karakteristik tersusun sebagai sembilan doblet di sekeliling dua singlet. Flagela dapat memiliki rambut, atau mastigonem, dan sisik yang menghubungkan membran dan batang internal. Interiornya menyatu dengan sitoplasma sel.
Reproduksi eukariota dilakukan melalui pembelahan sel, yang umumnya terjadi secara mitosis, yaitu proses pembelahan inti sel yang menyebabkan sebuah sel anak menerima duplikat setiap kromosom yang dimiliki sel induk. Pada kebanyakan eukariota terdapat juga reproduksi seksual, di antara sel haploid, yaitu sel yang hanya memiliki satu buah kromosom dari masing-masing pasang kromosom yang dimiliki sel induk yang melibatkan proses fusi inti sel (singami) dan pembelahan secara meiosis yang menghasilkan sel diploid, yaitu sel yang memiliki pasangan kromosom yang lengkap.
Eukariota adalah organisme dengan sel yang memiliki nukleus dan organel bermembran lainnya.
Eukariota berada dalam takson Eukarya atau Eukaryota. Hal yang membuat sel eukariotik berbeda dari sel prokariotik (Bacteria dan Archaea) adalah bahwa mereka memiliki organel bermembran, terutama inti, yang berisi materi genetik, dan terbungkus oleh selubung nukleus. Nama Eukaryota digunakan karena adanya inti, yang berasal dari bahasa Yunani ευ (eu, "baik") dan κάρυον (karyon, "kacang" atau "kernel"). Sel eukariotik juga mengandung organel bermembran lainnya seperti mitokondria dan badan Golgi. Selain itu, tumbuhan dan alga mengandung kloroplas. Organisme eukariotik bisa berbentuk uniseluler atau multiseluler. Hanya eukariota yang memiliki banyak jenis jaringan yang terdiri dari jenis sel yang berbeda.
Eukariota dapat bereproduksi baik melalui reproduksi aseksual melalui mitosis dan reproduksi seksual melalui meiosis. Dalam mitosis, satu sel membelah untuk menghasilkan dua sel yang identik secara genetik. Dalam meiosis, replikasi DNA diikuti oleh dua putaran pembelahan sel untuk menghasilkan empat sel anak masing-masing dengan setengah jumlah kromosom dari sel induknya (sel haploid). Sel-sel ini bertindak sebagai sel kelamin (gamet – masing-masing gamet hanya memiliki satu pelengkap kromosom, masing-masing campuran unik dari pasangan kromosom orang tua yang sesuai) yang dihasilkan dari rekombinasi genetik selama meiosis.
Domain Eukaryota tampaknya monofiletik, dan membuat salah satu dari tiga domain kehidupan. Dua domain lainnya, Bacteria dan Archaea, adalah prokariota dan tidak memiliki ciri-ciri di atas. Eukariota merupakan minoritas kecil dari semua makhluk hidup. Namun, karena ukuran mereka jauh lebih besar, biomassa kolektif eukariota di seluruh dunia diperkirakan sekitar setara dengan prokariota. Eukariota pertama kali berkembang sekitar 1,6-2,1 miliar tahun yang lalu.
Heilkjörnungar (fræðiheiti: Eukaryota) eru lífverur með frumur þar sem frumukjarninn (eða -kjarnarnir) er hulinn frumuhimnu. Heilkjörnungar telja bæði dýr, jurtir og sveppi (sem eru flest fjölfruma) auk ýmissa annarra hópa sem stundum eru flokkaðir sem frumverur og eru margir einfruma. Hinn meginhópur lífvera er dreifkjörnungar sem ekki eru með aðgreindan kjarna eða önnur frumulíffæri og telja gerla og fornbakteríur. Heilkjörnungar eiga sér sameiginlegan uppruna og eru stundum skilgreindir sem veldi eða lén.
Heilkjörnungar (fræðiheiti: Eukaryota) eru lífverur með frumur þar sem frumukjarninn (eða -kjarnarnir) er hulinn frumuhimnu. Heilkjörnungar telja bæði dýr, jurtir og sveppi (sem eru flest fjölfruma) auk ýmissa annarra hópa sem stundum eru flokkaðir sem frumverur og eru margir einfruma. Hinn meginhópur lífvera er dreifkjörnungar sem ekki eru með aðgreindan kjarna eða önnur frumulíffæri og telja gerla og fornbakteríur. Heilkjörnungar eiga sér sameiginlegan uppruna og eru stundum skilgreindir sem veldi eða lén.
Gli eucarioti (Eukaryota, dal greco εὖ eu «buono» e κάρυον káryon «nucleo»[1]) sono uno dei due domini della classificazione tassonomica degli esseri viventi. Costituiscono il dominio più complesso e includono cinque regni: piante, funghi, animali, protisti e cromisti.
Il criterio per la distinzione dall'altro dominio, i procarioti, è la presenza di un nucleo cellulare ben definito, isolato dal resto della cellula tramite una membrana e contenente la maggior parte del materiale genetico rappresentato dal DNA (una parte minore è contenuta nei mitocondri). Gli eucarioti possiedono inoltre un sistema endomembranoso.
Gli eucarioti si distinguono dai procarioti anche per numerose caratteristiche a livello molecolare. Ad esempio:
Solo negli eucarioti si ha riproduzione sessuale, che presenta due modi di divisione: la mitosi e la meiosi. La mitosi è il processo di divisione che serve principalmente all'accrescimento e al rinnovo dei tessuti, mentre la meiosi è specifica delle cellule adibite alla riproduzione, dette gameti. Mediante mitosi una cellula fa una copia di sé stessa con lo stesso numero di cromosomi, formando così due cellule diploidi. La meiosi si svolge invece in due sequenze, in cui una cellula diploide, con un metodo caratteristico, si sdoppia in due cellule aploidi (ovvero con una sola copia di ciascun cromosoma) e poi, durante la seconda sequenza, queste due cellule si dividono formando alla fine quattro cellule aploidi.
La transizione dai procarioti agli eucarioti ha rappresentato per molti studiosi uno dei passaggi evolutivi più importanti, secondo solo a quello dell'evoluzione delle cellule fotosintetiche. Il problema di come possa essere avvenuto questo passaggio è stato argomento di accese discussioni. Secondo l'ipotesi più diffusa, per circa 2 miliardi di anni, quindi per un tempo maggiore della metà di quello trascorso dall'inizio della vita, sono esistite solo cellule prokaryota.[3] L'origine della cellula eucariota risalirebbe all'incirca a 1,5 miliardi di anni fa in pieno Precambriano, quando alcuni procarioti si stabilirono all'interno di altri organismi in una sorta di "simbiosi interna permanente". Esistono sufficienti prove che gli eucarioti derivano dai procarioti attraverso tale meccanismo di endosimbiosi (Serial Endosymbiosis Theory), ipotizzato in forma completa da Lynn Margulis negli anni sessanta del XX secolo.
Questa origine può essere distinta in due tappe:
Secondo una tassonomia consolidata (anche se ormai obsoleta per vari aspetti), gli eucarioti pluricellulari venivano ripartiti fra i tre regni delle Piante, dei Funghi e degli Animali, mentre quelli unicellulari venivano di solito riuniti nel regno dei Protisti (Robert Whittaker 1969, Carl Woese 1977).
Successivamente le alghe unicellulari sono state incluse dalla maggioranza degli studiosi nel regno delle Piante e i rimanenti organismi unicellulari sono stati divisi da alcuni scienziati in regni separati: (Chromista e Protozoa secondo Thomas Cavalier-Smith).
Recentemente (per es. Sina et al., 2005[4] Keeling et al., 2005), sulla base di caratteri strutturali e molecolari, è stato proposto di dividere gli organismi eucarioti in sei grandi gruppi (chiamati supergruppi) accettati universalmente, ma le cui relazioni e la cui monofilia non sono ancora completamente chiarite:
Un'ulteriore proposta di classificazione[5] suggerisce che gli organismi eucarioti possano venir suddivisi in due soli grandi cladi sulla base della loro derivazione da organismi ancestrali con uno o due flagelli:
Un'ulteriore proposta considera invece:
Alghe rosse (Rhodophyta) ![]()
Piante verdi (Viridiplantae) .jpg)
Discoba (Excavata) _(cropped).jpg)
Holomycota (inc. funghi) ![]()
Holozoa (inc. animali) ![]()
Gli eucarioti (Eukaryota, dal greco εὖ eu «buono» e κάρυον káryon «nucleo») sono uno dei due domini della classificazione tassonomica degli esseri viventi. Costituiscono il dominio più complesso e includono cinque regni: piante, funghi, animali, protisti e cromisti.
Il criterio per la distinzione dall'altro dominio, i procarioti, è la presenza di un nucleo cellulare ben definito, isolato dal resto della cellula tramite una membrana e contenente la maggior parte del materiale genetico rappresentato dal DNA (una parte minore è contenuta nei mitocondri). Gli eucarioti possiedono inoltre un sistema endomembranoso.
Eukaryota (nomen a Whittaker & Margulis anno 1978 a verbis Graecis creatum: εὐ- 'bonus, bene' et κάρυον 'nucleus') sunt omnia vitalia quae nucleum, mitochondria, et organella in suis cellulis possident. Et regnum animalium et regnum plantarum sunt eukaryota.
Lichenes sunt casus specialis, quia et ex fungis et ex algis constituunt.
Casus specialis et Myxomycetes sunt quia cellulas maximas cum multis nucleis formant. Cellulae non solent vallum habere, sed si fructuant, myxomycetes cellulas cum vallo formant.
Eukaryota (nomen a Whittaker & Margulis anno 1978 a verbis Graecis creatum: εὐ- 'bonus, bene' et κάρυον 'nucleus') sunt omnia vitalia quae nucleum, mitochondria, et organella in suis cellulis possident. Et regnum animalium et regnum plantarum sunt eukaryota.
Lichenes sunt casus specialis, quia et ex fungis et ex algis constituunt.
Casus specialis et Myxomycetes sunt quia cellulas maximas cum multis nucleis formant. Cellulae non solent vallum habere, sed si fructuant, myxomycetes cellulas cum vallo formant.
Eukariotinė ląstelė (sen. gr. eu – tikras, karyon – branduolys) – tai ląstelės, turinčios tikrą branduolį, kurį nuo citoplazmos atskiria membrana. Ląstelių dydis apie 13 mikrometrų.
Citoplazmoje išsidėstę įvairūs organoidai, kuriuos galima pamatyti tik pro mikroskopą, o kai kuriuos tik pro elektroninį. Ląstelės organoidai nuo citoplazmos atskirti:
Kai kurios eukariotinės ląstelės turi išaugas – žiuželius, judėjimo skystyje organeles. Be žiuželių būna fimbrijos (blakstienėlės) – trumpos ataugos, padedančios prilipti prie ląstelių ar kitų daiktų.
Yra trys pagrindinės eukariotinių ląstelių formos:
Augalų ląstelės Grybų ląstelės Gyvūnų ląstelės Ląstelės sienelė Iš celiuliozės Daugiausia iš chitino Nėra Centrinė vakuolė Yra Yra Nėra Plastidės Yra Nėra Nėra Tipiškas rezervinis angliavandenis Krakmolas Glikogenas Glikogenas Centriolė Būna retai Būna retai Yra
Eikarioti (Eukaryota vai Eukarya) ir organismi, kas sastāv no šūnām, kurām ir morfoloģiski labi izveidoti šūnas kodoli un ar membrānām klāti organoīdi. Eikarioti ir viena no organismu impērijām. Eikariotu impērijā ietilpst augi, dzīvnieki, sēnes un protisti. Organismus ar pretējām īpašībām — bez kodola un bez citiem šūnas organoīdiem, kam ir savas membrānas — sauc par prokariotiem (pie prokariotiem pieder baktērijas un arheji). Vārds ‘eikariots’ ir radies no grieķu valodas, kur εὖ (eû) nozīmē ‘īsts’, bet κάρυον (káruon) — ‘kodols’.
Eikarioti vairojas gan bezdzimumvairošanās ceļā mitozes procesā, gan dzimumvairošanās ceļā mejozes procesā.
Eikariotu sugu skaits ir pavisam neliels, salīdzinot ar prokariotu sugu skaitu.[1] Toties, tā kā eikarioti ir daudz lielāki un sastāv no daudzām šūnām, tad eikariotu biomasa pasaulē ir aptuveni tikpat liela kā prokariotu biomasa.[1] Eikarioti uz Zemes radās aptuveni pirms 1,6–2,1 miljardiem gadu.
Eikariotu šūnas ir daudz lielākas par prokariotu šūnām, parasti to lielumus pārsniedz pat tūkstošiem reižu. Šūnas kodols norobežots ar divām membrānām. Šūnām ir iekšējo organoīdu daudzveidība un citoskelets, kas veidots no mikrocaurulītēm un mikrofilamentiem, kuri ieņem nozīmīgu lomu šūnas organizēšanā un tās uzbūvē. Organoīdi ir klāti ar membrānām. Eikariotu šūnu kodolos atrodas hromatīns — dezoksiribonukleīnskābes (DNS) pavedieni, kas šūnu dalīšanās laikā atrodas pavedienveida struktūrās, kuras sauc par hromosomām.
Starp eikariotu šūnām pastāv arī atšķirības. Piemēram, augu šūnās ir hloroplasti, kas nav citu eikariotu šūnās. Augu šūnās ir arī celulozes šūnapvalks, ar šūnsulu pildītas vakuolas un plastīdas. Savukārt vienīgi dzīvnieku šūnās ir centrosoma, kuru veido divas centriolas.
Sēņu šūnās nav plastīdas un ar šūnsulu pildītas vakuolas. Šūnapvalks satur hitīnu. Šūnās atrodas hifas — mikroskopiski šūnu pavedieni, kas veido micēliju.
Vēsturiski eikariotus dalīja divās grupās — augos un dzīvniekos. Viens no pirmajiem, kas šādu iedalījumu piedāvāja, bija zviedru botāniķis un zoologs Kārlis Linnejs. Arī vienšūnas eikariotus, kad tie tika atklāti, ieskaitīja vai nu pie augiem, vai pie dzīvniekiem. 1866. gadā vācu biologs Ernsts Hekels izdalīja atsevišķu organismu valsti — protistus. Tajā ietilpa ne tikai vienšūņi, bet arī aļģes. Visbeidzot no augiem atsevišķi tika atdalīta sēņu valsts.
Mūsdienās eikariotu impērija dalās četrās organismu valstīs:
Pēc jaunākiem DNS pētījumiem tiek pārskatīts eikariotu iedalījums. Pastāv dažādi jauni eikariotu klasifikācijas piedāvājumi. Viens no dominējošajiem ir eikariotu iedalījums piecās virsgrupās: Archaeplastida, SAR virsgrupa, Excavate, Amoebozoa un Opisthokonta.
Eikarioti (Eukaryota vai Eukarya) ir organismi, kas sastāv no šūnām, kurām ir morfoloģiski labi izveidoti šūnas kodoli un ar membrānām klāti organoīdi. Eikarioti ir viena no organismu impērijām. Eikariotu impērijā ietilpst augi, dzīvnieki, sēnes un protisti. Organismus ar pretējām īpašībām — bez kodola un bez citiem šūnas organoīdiem, kam ir savas membrānas — sauc par prokariotiem (pie prokariotiem pieder baktērijas un arheji). Vārds ‘eikariots’ ir radies no grieķu valodas, kur εὖ (eû) nozīmē ‘īsts’, bet κάρυον (káruon) — ‘kodols’.
Eikarioti vairojas gan bezdzimumvairošanās ceļā mitozes procesā, gan dzimumvairošanās ceļā mejozes procesā.
Eikariotu sugu skaits ir pavisam neliels, salīdzinot ar prokariotu sugu skaitu. Toties, tā kā eikarioti ir daudz lielāki un sastāv no daudzām šūnām, tad eikariotu biomasa pasaulē ir aptuveni tikpat liela kā prokariotu biomasa. Eikarioti uz Zemes radās aptuveni pirms 1,6–2,1 miljardiem gadu.
Eukariot ialah sejenis organisma yang sel-selnya mengandungi struktur-struktur kompleks yang terlindung dalam membran. Eukariot dikenali secara formal dengan nama takson Eukarya atau Eukaryota. Struktur ikatan membran yang membezakan sel eukariot daripada sel prokariot ialah nukleus, atau sampul nukleus yang mengandungi bahan genetik.[1][2][3] Eukariot dinamai sedemikian kerana kehadiran nukleus, iaitu gabungan perkataan Yunani ευ (eu, "baik") dan κάρυον (karyon, "kacang" atau "inti"). Kebanyakan sel eukariot juga mengandungi lain-lain organel ikatan membran seperti mitokondria, kloroplas dan radas Golgi. Semua spesies organisme eukariot yang besar dan kompleks merupakan eukariot, termasuk haiwan, tumbuhan dan kulat, namun kebanyakan spesies eukariot merupakan mikroorganisma protis.
Pembahagian sel dalam eukariot adalah berbeza dengan organisma tanpa nukelus (prokariot). Ia melibatkan pengasingan kromosom yang diduakan melalui pergerakan yang diarahkan oleh mikrotubul. Terdapat dua jenis proses pembahagian, iaitu mitosis yang mana satu sel berpecah kepada dua sel yang seiras secara genetik; dan meiosis yang diperlukan dalam pembiakan seks, iaitu satu sel diploid (dengan dua kromosom, satu dari setiap induk) menjalani pencantuman semula setiap pasang kromosom induk, kemudian dua peringkat pembahagian sel, maka terhasilnya empat sel haploid (gamet). Setiap gamet mempunyai hanya satu pelengkap kromosom, yang mana setiap satu kromosom ini merupakan campuran unik pasangan kromosom induknya.
Eukariot kelihatan sebagai monofiletik, dan oleh itu merupakan salah satu daripada tiga domain hidupan. Dua lagi domain, iaitu Bacteria dan Archaea, merupakan prokariot dan tidak memiliki ciri-ciri yang disebut di atas. Eukariot merupakan minoriti yang sangat sedikit di kalangan segala hidupan; dalam tubuh manusia pun terdapat 10 kali lebih banyak mikroogranisma berbanding sel manusia.[4] Namun demikian, disebabkan saiznya yang lebih besar, jumlah terkumpul biojisim sedunia eukariot dianggarkan lebih kurang setara dengan prokariot.[5]
Rencana ini mengandungi bahan-bahan daripada Buku Teks Peringkat Permulaan Sains yang diterbitkan oleh Pusat Maklumat Bioteknologi Nasional (NCBI). Sebagai terbitan kerajaan Amerika Syarikat, buku ini berada dalam domain awam.
Eukariot ialah sejenis organisma yang sel-selnya mengandungi struktur-struktur kompleks yang terlindung dalam membran. Eukariot dikenali secara formal dengan nama takson Eukarya atau Eukaryota. Struktur ikatan membran yang membezakan sel eukariot daripada sel prokariot ialah nukleus, atau sampul nukleus yang mengandungi bahan genetik. Eukariot dinamai sedemikian kerana kehadiran nukleus, iaitu gabungan perkataan Yunani ευ (eu, "baik") dan κάρυον (karyon, "kacang" atau "inti"). Kebanyakan sel eukariot juga mengandungi lain-lain organel ikatan membran seperti mitokondria, kloroplas dan radas Golgi. Semua spesies organisme eukariot yang besar dan kompleks merupakan eukariot, termasuk haiwan, tumbuhan dan kulat, namun kebanyakan spesies eukariot merupakan mikroorganisma protis.
Pembahagian sel dalam eukariot adalah berbeza dengan organisma tanpa nukelus (prokariot). Ia melibatkan pengasingan kromosom yang diduakan melalui pergerakan yang diarahkan oleh mikrotubul. Terdapat dua jenis proses pembahagian, iaitu mitosis yang mana satu sel berpecah kepada dua sel yang seiras secara genetik; dan meiosis yang diperlukan dalam pembiakan seks, iaitu satu sel diploid (dengan dua kromosom, satu dari setiap induk) menjalani pencantuman semula setiap pasang kromosom induk, kemudian dua peringkat pembahagian sel, maka terhasilnya empat sel haploid (gamet). Setiap gamet mempunyai hanya satu pelengkap kromosom, yang mana setiap satu kromosom ini merupakan campuran unik pasangan kromosom induknya.
Eukariot kelihatan sebagai monofiletik, dan oleh itu merupakan salah satu daripada tiga domain hidupan. Dua lagi domain, iaitu Bacteria dan Archaea, merupakan prokariot dan tidak memiliki ciri-ciri yang disebut di atas. Eukariot merupakan minoriti yang sangat sedikit di kalangan segala hidupan; dalam tubuh manusia pun terdapat 10 kali lebih banyak mikroogranisma berbanding sel manusia. Namun demikian, disebabkan saiznya yang lebih besar, jumlah terkumpul biojisim sedunia eukariot dianggarkan lebih kurang setara dengan prokariot.
Eukaryoten, ook wel Eucarya,[2] (van Grieks: εὖ = echt, geheel; κάρυον = kern; volledige kern) vormen een van de drie domeinen waarin het leven verdeeld kan worden. Het zijn alle organismen waarvan de cellen minstens één celkern bevatten. Naast een celkern hebben de meeste eukaryotische cellen ook andere organellen, zoals mitochondriën, chloroplasten en golgiapparaat. De term "eukaryoot" (of "eukaryotisch") wordt ook als bijvoeglijk naamwoord gebruikt en duidt op de aanwezigheid van een kern.
De andere twee, prokaryotische, domeinen, de Bacteria en Archaea, werden eerder samengenomen als éen domein, dat van de Monera. Tegenwoordig worden de Archaea beschouwd als de zustergroep van de Eukaryota.
Prokaryoten hebben een celbouw waarin het DNA niet omsloten wordt door een kernmembraan maar vrij in het cytoplasma ligt. Andere celorganellen ontbreken hier ook.
De protisten, alle eencellige, eukaryotische micro-organismen, vormen binnen het domein van de eukaryoten de grootste, onderling grotendeels weinig verwante groep, met de grootste soortenrijkdom.
Vrijwel alle grotere, meercellige organismen, zoals alle planten (in de supergroep Archaeplastida), dieren en schimmels (beide in de supergroep Unikonta), zijn eveneens eukaryoten. Ook algen als roodwieren en bruinwieren zijn veelal meercellig en kunnen soms zeer groot worden.
Eukaryoten zijn verschillend van de prokaryoten, de organismen in de bacteriële domeinen. Eukaryoten hebben interne structuren die gebonden zijn aan membranen, zoals de celkern (die de minimumvoorwaarde vormt voor een eukaryoot), ze hebben een cytoskelet van microtubuli en ze hebben kenmerkende ribosomen.
Als men de prokaryoten en de eukaryoten met elkaar vergelijkt, dan lijken de eukaryoten chimerische organismen. Evolutie heeft dan ook niet alleen langs Darwiniaanse lijnen (zoals mutaties en selectie) plaatsgevonden, maar waarschijnlijk ook langs niet-Darwiniaanse lijnen als endosymbiose en endosymbiotische genoverdracht (EGT), versmelting van cellen en horizontale genoverdracht (HGT). Door een dergelijke constructiewijze moeten bij de soortvorming de resulterende veranderingen geleid hebben tot grote evolutionaire sprongen en niet tot een geleidelijke evolutie. De grootste overgang is die van prokaryoten (waarschijnlijk een voorouder van de tegenwoordige Archaea) naar de eukaryoten.
Eukaryotische cellen zijn meestal veel groter dan prokaryotische. Ze hebben, in tegenstelling tot de laatsten, verschillende interne membranen en structuren, die organellen genoemd worden. Eukaryote cellen hebben cytoplasma en een cytoskelet dat is opgebouwd uit microtubuli, microfilamenten en intermediaire filamenten, die de interne organisatie en vorm van de cel bepalen. Ze bevatten een uitwendig membraan waarin het cytoplasma ligt, een plasma waarin de belangrijkste organellen drijven.
De celkern is omgeven door een dubbel membraan met kernporiën waardoor materiaal aan- en afgevoerd kan worden. Het DNA van een eukaryotische cel ligt in de celkern, waar het verdeeld is in chromosomen. Dit membraan wordt omgeven door een ingewikkeld netwerk van ronde of vlakke uitstulpingen, die samen het endoplasmatisch reticulum heten. Het endoplasmatisch reticulum kan worden verdeeld in een ruw reticulum dat zich direct om de celkern heen bevindt, en een glad reticulum verder van de kern af. Het ruwe gedeelte herbergt ribosomen, organellen die eiwitten aanmaken. Deze eiwitten worden door het reticulum naar buiten getransporteerd. In het gladde reticulum worden ze omgeven door vesikels die van het reticulum afsplitsen.
In het golgicomplex worden de producten afkomstig van het endoplasmatisch reticulum (ER) omgebouwd en opgeslagen, om dan later naar andere bestemmingen getransporteerd te worden. Bij de meeste eukaryoten bestaat het uit afgeplatte en opgestapelde vesikels. Zulke stapels eiwithoudende vesikels worden golgiapparaat of dictyosoom genoemd.
Eukaryotische cellen bevatten door membranen gevormde structuren, die samen het endomembraansysteem genoemd worden. Door afsplitsingen van de membranen kunnen soms blaasjes ontstaan, die vesikels of vacuolen genoemd worden. Eukaryotische cellen nemen voedsel op door een proces dat endocytose genoemd wordt. Het membraan omsluit het op te nemen voorwerp en splitst ten slotte, zodat zich een vesikel rondom de stof vormt. Vesikels kunnen verschillende functies binnenin de cel hebben, zoals:
Bij planten wordt het grootste deel van het volume van uitgegroeide cellen opgenomen door de vacuole, dat dient om de turgordruk op peil te houden.
Bijna alle eukaryoten hebben mitochondriën voor hun energievoorziening, die vrij in het cytoplasma liggen. Mitochondriën worden omhuld door een dubbele membraan van fosfolipiden. Het binnenste membraan is in kleine instulpingen naar binnen geplooid waar respiratie plaatsvindt, deze worden cristae genoemd. Mitochondrien bevatten hun eigen DNA (het mtDNA) en ribosomen en zorgen voor hun eigen reproductie. Het aantal mitochondriën in een eukaryotische cel hangt af van de energiebehoefte. Een klein aantal eukaryoten heeft geen mitochondriën, maar uit mitochondriën afgeleide organellen die een vergelijkbare functie hebben, zoals hydrogenosomen of mitosomen.
Bij de planten en verschillende groepen algen komen daarnaast organellen voor die plastiden genoemd worden. Net als mitochondriën bevatten deze hun eigen DNA, het plastoom. Er zijn verschillende typen plastiden, maar de belangrijkste soort zijn chloroplasten. Chloroplasten bevatten het biopigment chlorofyl, waarmee ze licht opvangen, waardoor ze voor fotosynthese zorgen. Chloroplasten spelen daarmee een belangrijke rol in de energievoorziening van de plantaardige cel. Andere soorten plastiden worden gebruikt om voedsel op te slaan in de vorm van bijvoorbeeld zetmeel.
Het cytoskelet bestaat uit verschillende soorten polymeren van eiwitten in de cellen, die samen zorgen voor stevigheid, vorm en beweeglijkheid. Het cytoskelet bestaat uit drie typen filamenten, namelijk microfilamenten, microtubuli en intermediaire filamenten.
Eukaryotische cellen kunnen aan de buitenkant van de celstructuren in het cytoskelet hebben die beweging mogelijk maken, zoals een of meerdere flagel of "zweepstaartjes", cilia of lamellipodia.
Flagellen zijn lange staartvormige structuren, ciliën zijn korter. Ze bestaan voornamelijk uit tubuline en kunnen gebruikt worden bij de voortbeweging, bij het binnenhalen van voedsel of bij aftasten van de omgeving. De flagellen van eurkaryoten verschillen sterk van die van prokaryoten. De flagellen worden ondersteund door een bundel microtubuli, typisch negen paren om twee enkele, die uit een centriool komen. Flagellen kunnen bedekt zijn met haarachtige structuren die mastigonemen genoemd worden, of schalen die membranen en interne structuren afdekken. Het binnenste van een flagel is verbonden met het cytoplasma van de cel.
Centriolen komen ook voor op cellen die geen flagella hebben. Vaak vormen ze de aanhechtingsplek van microtubuli op de cel. Deze structuren in het cytoskelet spelen een rol tijdens de celdeling. Flagellen en ciliën kunnen reageren op chemische signalen door beweging van de cel te veroorzaken, zoals bij chemotaxis of chemokinese.
Sommige protisten, zoals Wortelpotigen, Stralendiertjes of Zonnediertjes, hebben lang uitgerekte structuren in het cytoskelet, die pseudopodia of schijnvoetjes genoemd worden. Ze kunnen dienen om deze eencellige organisme in water te laten drijven of een prooi te vangen. Haptophyta hebben een uniek organel, een haptonema, dat lijkt op een flagel maar een andere opbouw van de microtubuli heeft.
In tegenstelling tot prokaryoten komt bij eukaryoten naast ongeslachtelijke voortplanting ook geslachtelijke voortplanting voor. Sommige groepen eukaryoten planten zich uitsluitend geslachtelijk voort, andere alleen op ongeslachtelijke wijze, en sommige kunnen zich op beide wijzen voortplanten en hebben een levenscyclus, soms met generatiewisseling.
De deling van de cel en de celkern gaat tegelijk, volgens een proces dat mitose genoemd wordt. Van elk chromosoom wordt hierbij een kopie gemaakt voor de dochtercel. De mitochondriën en plastiden reproduceren zich onafhankelijk door binaire deling en worden geërfd via de eicel.
In eukaryotische diploïde cellen zijn van elk chromosoom twee kopieën aanwezig. Bij geslachtelijke voortplanting worden haploïde geslachtscellen (gameten) aangemaakt door een proces dat meiose wordt genoemd. Dit zijn cellen waarin niet twee, maar één kopie aanwezig is van elk chromosoom. Bij de bevruchting fuseren twee haploïde cellen, van elke ouder één, zodat seksueel voortplantende eukaryoten twee ouders hebben. De manier waarop de meiose en celfusie plaatsvinden verschilt echter sterk tussen de verschillende groepen eukaryoten.
Omdat eukaryotische cellen groter zijn dan prokaryotische, hebben ze een relatief klein oppervlak en daarom een tragere stofwisseling. Ook zit er daarom meer tijd tussen twee celdelingen.
Het domein Eukaryoten wordt tegenwoordig onderverdeeld in vijf (of soms zes) supergroepen:
Haeckel (1894)De Franse bioloog Édouard Chatton (1883-1947) was in 1937 de eerste bioloog die cellen in twee typen verdeelde: eukaryoten en prokaryoten. In 1990 werd door Carl Woese en collega's het drie domeinensysteem ingevoerd, dat de oudere verdeling in vijf rijken verving. De Eukaryota vormen een van de drie domeinen van de moderne taxonomie, hoewel de vijf rijken soms nog op de middelbare scholen en andere niet-wetenschappelijke instellingen worden gebruikt. De verdeling in drie domeinen is gebaseerd op moleculair-genetisch onderzoek, waarin door vergelijkingen tussen DNA en RNA de evolutionaire verwantschappen tussen organismen worden bepaald.
Het DNA in de eukaryotische celkern bevat meer overeenkomsten met de Archaea dan met de Bacteria, zodat eukaryoten en Archaea weleens samen in een bovenliggend clade Neomura geplaatst worden. In andere opzichten, zoals het celmembraan, lijken eukaryoten echter weer sterker op de bacteriën.
De Eukaryoten werden in het vijf rijkensysteem onderverdeeld in vier rijken:
De protisten bleken later een parafyletische groep te zijn. In de moderne indeling worden de eukaryoten in zes supergroepen verdeeld (tussen het niveau van de Rijken en Domeinen):[3][4][5]
De eerste twee supergroepen vallen samen in de clade Unikonta, terwijl de overige groepen in de clade Bikonta vallen. De eerste groep heeft cellen die slechts een enkele flagel hebben, de tweede groep heeft twee of meer flagellen.[6] De verdere onderverdeling wordt voortdurend aangepast aan nieuwe inzichten, zo zijn voor de Archaeplastida en Chromalveolata aanwijzingen gevonden dat het geen monofyletische groepen zijn.[7]
Hieronder staan twee van de mogelijke cladogrammen met de positie van het domein van de Eukarya, van de zustergroepen en de afstammende supergroepen.[8]
Een alternatieve indeling is:
LUCABacteria (bacteriën)
Archaea ("oerbacteriën")
Archaeplastida (Primoplantae)
Onder de oudste fossielen van eukaryoten bevinden zich 1650 miljoen jaar oude Acritarcha. Nog ouder (2100 miljoen jaar) zijn fossielen van Grypania, een organisme waarvan vanwege zijn grootte wordt vermoed dat het een (eukaryotische) soort alg was. De oudste fossiele eukaryoten die aan een modern taxon kunnen worden toegeschreven zijn van de roodalg Bangiomorpha pubescens.[9] Er zijn zelfs chemische stoffen in 2,7 miljard jaar oud gesteente gevonden die typisch zijn voor activiteit van eukaryotisch leven,[10] maar wanneer de eerste eukaryoten ook verschenen, de fossielen laten duidelijk zien dat ze pas rond 1,2 miljard jaar geleden begonnen te diversificeren.
De eerste eukaryoten moeten uit prokaryotische voorouders ontstaan zijn, dankzij een proces dat endosymbiose genoemd wordt. De mitochondriën in eukaryotische cellen stammen volgens deze hypothese af van aerobe bacteriën die een andere, grotere cel binnendrongen en binnenin een beschermde omgeving vonden. De gastcel profiteerde op zijn beurt van de extra energiebron. De bacteriën verloren vervolgens overbodige delen van hun DNA en werden volledig afhankelijk van de gastheer, die op deze wijze een eukaryoot was geworden. Op vergelijkbare manier wordt verondersteld dat de eerste plastiden fotosynthetiserende blauwalgen waren, die een eukaryotische cel binnendrongen.[11]
Eukaryoten, ook wel Eucarya, (van Grieks: εὖ = echt, geheel; κάρυον = kern; volledige kern) vormen een van de drie domeinen waarin het leven verdeeld kan worden. Het zijn alle organismen waarvan de cellen minstens één celkern bevatten. Naast een celkern hebben de meeste eukaryotische cellen ook andere organellen, zoals mitochondriën, chloroplasten en golgiapparaat. De term "eukaryoot" (of "eukaryotisch") wordt ook als bijvoeglijk naamwoord gebruikt en duidt op de aanwezigheid van een kern.
De andere twee, prokaryotische, domeinen, de Bacteria en Archaea, werden eerder samengenomen als éen domein, dat van de Monera. Tegenwoordig worden de Archaea beschouwd als de zustergroep van de Eukaryota.
Prokaryoten hebben een celbouw waarin het DNA niet omsloten wordt door een kernmembraan maar vrij in het cytoplasma ligt. Andere celorganellen ontbreken hier ook.
De protisten, alle eencellige, eukaryotische micro-organismen, vormen binnen het domein van de eukaryoten de grootste, onderling grotendeels weinig verwante groep, met de grootste soortenrijkdom.
Vrijwel alle grotere, meercellige organismen, zoals alle planten (in de supergroep Archaeplastida), dieren en schimmels (beide in de supergroep Unikonta), zijn eveneens eukaryoten. Ook algen als roodwieren en bruinwieren zijn veelal meercellig en kunnen soms zeer groot worden.
Eukaryotar er éin- eller fleircella organismar kjenneteikna av indre membranar, eit celleskjelett og mange celleorganeller som ikkje finst i prokaryote celler. Dei viktigaste eukarote celleorganella er cellekjernen (nukleus), endoplasmatisk retikulum, golgiapparatet, celleskjelettet, mitokondriar, kloroplastar og flimmerhår (ciliar).
Domenet har fått namnet sitt etter den tydelege cellekjernen gjennom samansettinga av gresk ευ, 'god, ekte' og κάρυον, 'nøtt'. Inni kjernen finn ein arvematerialet DNA ordna i kromosom. I prokaryotar ligg derimot arvematerialet spreidd rundt i cella.
Eukaryotar er éin- eller fleircella organismar kjenneteikna av indre membranar, eit celleskjelett og mange celleorganeller som ikkje finst i prokaryote celler. Dei viktigaste eukarote celleorganella er cellekjernen (nukleus), endoplasmatisk retikulum, golgiapparatet, celleskjelettet, mitokondriar, kloroplastar og flimmerhår (ciliar).
Domenet har fått namnet sitt etter den tydelege cellekjernen gjennom samansettinga av gresk ευ, 'god, ekte' og κάρυον, 'nøtt'. Inni kjernen finn ein arvematerialet DNA ordna i kromosom. I prokaryotar ligg derimot arvematerialet spreidd rundt i cella.
Eukaryoter er én- eller flercellede organismer som er kjennetegnet ved mange celleorganeller som ikke forekommer i prokaryote celler. De viktigste eukarote celleorganellene er cellekjernen (nukleus), endoplasmatisk retikulum, golgiapparatet, celleskjelettet, mitokondrier, kloroplaster og flimmerhår (cilier).
Av disse celleorganellene forekommer cellekjerner, endoplasmatisk retikulum, golgiapparat og celleskjelett hos samtlige eukaryoter. Kloroplaster har med sikkerhet ikke vært til stede i eukaryotenes siste felles stamart. Hvordan det forholder seg med mitokondrier og flimmerhår, er fortsatt et åpent spørsmål. Det fins noen få eukaryote organismegrupper som mangler én eller begge av disse organellene (bl.a. Tetramastigota og Microsporidea). Tidligere antok man at dette tydet på at de respektive gruppene har overlevd fra en tid da opprinnelige eukaryoter fremdeles manglet disse organellene. Nå går man derimot ut ifra at disse gruppene ble sekundært forenklet, at de altså stammer fra organismer som har hatt både mitokondrier og flimmerhår. Diameteren på en eukaryot celle ligger stort sett mellom 1 og 100 µm.
Det har eksistert eukaryote organismer i mer enn to milliarder år. Deres siste felles stamart var éncellet og hadde altså etter alt å dømme evolvert en cellekjerne og et par med flimmerhår. Videre hadde den tatt opp en purpurbakterie, som etter hvert ble til den organellen som man i dag kjenner som mitokondrium. Ved den samme mekanismen opptak av frittlevende organismer (endosymbiose) oppstod senere, og flere ganger uavhengig, kloroplaster. I løpet av årmilliardene har denne gruppen fått flere og flere arter, deriblant mange nokså komplekse og mange flercellede.
Av de cirka 1,8 millioner arter av eukaryoter som er kjent i dag, er hovedparten flercellet (hovedsakelig dyr og planter). Men dette skyldes sannsynligvis at de flercellede gruppene er best undersøkt. Man går ut ifra at antallet arter som eksisterer, er minst det hundredobbelte av de som er beskrevet så langt, og at encellede organismer kan være i flertall blant disse. Det biologiske mangfoldet er altså ikke mindre blant de encellede artene verken morfologisk, økologisk eller taksonomisk.
Eukaryoter regnes innenfor tradisjonell systematikk som en av tre hovedgrupper av levende organismer, hvor bakterier og arkebakterier utgjør de to første, og den siste gruppen av eukaryoter siden gir opphav til protister, planter, sopp og dyr.[1] I tradisjonell, men omstridt systematikk er disse fem-seks gruppene ofte omtalt som «rikene» av liv innenfor biologien. det gir eukaryotene status som opphav for fire distinkte «riker» eller hovedgrupper i systematikken:[2]
Det fins mange måter å dele inn de eukaryote organismene på. De fleste av disse forteller imidlertid veldig lite om gruppenes innbyrdes slektskapsforhold. Én mulig inndeling, som for tiden er nokså populær i lærebøker, er i fire «riker», de tre flercellede gruppene dyr, planter og sopp samt en overveiende encellet gruppe, protoktister («Protoctista»).
I denne inndelingen er den sistnevnte gruppen en håpløst parafyletisk gruppe, fordi mange protoktister er i nærmere slekt med enten dyr, planter eller sopp enn med andre protoktister. Flercellethet har oppstått flere ganger uavhengig i veldig ulike grupper av encellede organismer. Også blant protokistene finner man dessuten mange former for og overganger til flercellethet, fra enkle «celleklumper» via mer komplekse kolonier med arbeidsdeling til meterstore thalli.
Problemet med evolusjonær slektskapsrekonstruksjon mellom eukaryoter er at mange av de encellede organismene er forholdsvis enkelt bygd, slik at man har få egenskaper å rekonstruere slektskap med. Molekylære og genetiske metoder har revolusjonert vårt syn på eukaryotenes slektskapsforhold, men har langt fra klart å oppklare alle forhold.
Noen av de tradisjonelle gruppene fikk støtte gjennom nye forskning (f.eks. foraminiferer og radiolarier). Ellers har bildet imidlertid endret seg fullstendig. På den ene siden har det vist seg at mang en kjent gruppe ikke er annet enn et svært kunstig sammensurium. På den andre siden har man oppdaget nære slektskapsbånd mellom grupper man tidligere hadde ansett for å være nokså ulike. Eksempler på det førstnevnte, altså på grupper som ikke har fått støtte gjennom den evolusjonære slektskapsforskningen, er:
Utvalgte eksempler på uventede slektskapsbånd er:
Slik oppsto den morderne celle - artikkel fra forskning.no 24.5.03
Eukaryoter er én- eller flercellede organismer som er kjennetegnet ved mange celleorganeller som ikke forekommer i prokaryote celler. De viktigste eukarote celleorganellene er cellekjernen (nukleus), endoplasmatisk retikulum, golgiapparatet, celleskjelettet, mitokondrier, kloroplaster og flimmerhår (cilier).
Av disse celleorganellene forekommer cellekjerner, endoplasmatisk retikulum, golgiapparat og celleskjelett hos samtlige eukaryoter. Kloroplaster har med sikkerhet ikke vært til stede i eukaryotenes siste felles stamart. Hvordan det forholder seg med mitokondrier og flimmerhår, er fortsatt et åpent spørsmål. Det fins noen få eukaryote organismegrupper som mangler én eller begge av disse organellene (bl.a. Tetramastigota og Microsporidea). Tidligere antok man at dette tydet på at de respektive gruppene har overlevd fra en tid da opprinnelige eukaryoter fremdeles manglet disse organellene. Nå går man derimot ut ifra at disse gruppene ble sekundært forenklet, at de altså stammer fra organismer som har hatt både mitokondrier og flimmerhår. Diameteren på en eukaryot celle ligger stort sett mellom 1 og 100 µm.
Det har eksistert eukaryote organismer i mer enn to milliarder år. Deres siste felles stamart var éncellet og hadde altså etter alt å dømme evolvert en cellekjerne og et par med flimmerhår. Videre hadde den tatt opp en purpurbakterie, som etter hvert ble til den organellen som man i dag kjenner som mitokondrium. Ved den samme mekanismen opptak av frittlevende organismer (endosymbiose) oppstod senere, og flere ganger uavhengig, kloroplaster. I løpet av årmilliardene har denne gruppen fått flere og flere arter, deriblant mange nokså komplekse og mange flercellede.
Av de cirka 1,8 millioner arter av eukaryoter som er kjent i dag, er hovedparten flercellet (hovedsakelig dyr og planter). Men dette skyldes sannsynligvis at de flercellede gruppene er best undersøkt. Man går ut ifra at antallet arter som eksisterer, er minst det hundredobbelte av de som er beskrevet så langt, og at encellede organismer kan være i flertall blant disse. Det biologiske mangfoldet er altså ikke mindre blant de encellede artene verken morfologisk, økologisk eller taksonomisk.
Eukarionty, eukariota, eukarioty, organizmy eukariotyczne (Eukaryota, Eukarya), określane też jako jądrowce, jądrowe, organizmy jądrowe, karioty, kariota (Karyobionta) – organizmy zbudowane z komórek posiadających jądro komórkowe z chromosomami, co jest jednym z elementów odróżniających je od prokariontów. Jądro komórkowe odgraniczone jest od cytoplazmy podwójną błoną białkowo-lipidową. Nazwa naukowa pochodzi od greckich słów ευ ("dobry", "prawdziwy") i κάρυον ("orzech", "jądro").
W zależności od ujęcia systematycznego jądrowce są traktowane jako domena (cesarstwo, nadkrólestwo) lub królestwo. Najpopularniejszą z tych kategorii systematycznych jest obecnie domena. Do eukariotów zalicza się wszystkie organizmy komórkowe, z wyjątkiem bakterii i archeowców, a więc prokariotów. Szacunki dotyczące liczby gatunków w obrębie tej grupy wahają się pomiędzy 5 a 10 mln[1][2]. Według analiz przeprowadzonych przez naukowców z Census of Marine Life, opublikowanych w sierpniu 2011 roku, liczba ta wynosi 8,7 (±1,3) mln[3], z czego naukowo opisane zostały mniej niż dwa miliony[2].
Komórki eukariotyczne są z zasady znacznie większe od prokariotycznych. Choć zmienność rozmiarów i jednych i drugich jest bardzo duża (najmniejsze eukarioty mają rozmiar mniejszy niż 1 µm), typowe proporcje objętości są rzędu tysiąc do jednego.
Prawdopodobnie organizmy eukariotyczne wywodzą się z prokariotycznych archeonów[4] (a w każdym razie są z nimi spokrewnione bliżej niż z bakteriami[5]), jednak ich wczesne dzieje ewolucyjne są słabo znane. Uznaje się za możliwe, że po prokariotycznych jeszcze przodkach wczesne jądrowce odziedziczyły zdolność do ruchu ameboidalnego i fagocytozy, lecz znacznie ją udoskonaliły. Głównym jednak "wynalazkiem" była reorganizacja materiału genetycznego. Komórka eukariotyczna zawiera wielokrotnie więcej materiału genetycznego niż prokariotyczna, dzięki czemu jest w stanie produkować więcej typów białek i ma potencjalnie nieograniczone możliwości regulacji. Materiał genetyczny występuje tu w postaci wielu skupionych w jądrze chromosomów. Są to liniowe fragmenty DNA związanego z licznymi białkami, które chronią go, powielają i precyzyjnie sterują ekspresją. Komórki eukariotyczne posiadają wiele organelli – zarówno analogiczne do występujących u prokariotów np. rybosomy czy chromosomy (odpowiadające bakteryjnemu genoforowi), jak i takie, których u żadnych prokariotów nie ma. Do tych drugich należy retikulum endoplazmatyczne wraz z błoną jądrową oraz mitochondria i plastydy (np. chloroplasty). Przynajmniej te dwa ostatnie pochodzą ze stosunkowo późnej w skali ewolucyjnej endosymbiozy komórek wczesnych jądrowych z bakteriami i sinicami. Wreszcie ponad miliard lat temu komórki eukariotyczne wykształciły możliwość rozmnażania drogą płciową, co jeszcze bardziej zwiększyło ich możliwości ewolucyjne[6].
Najstarsze znane skamieniałości, które prawdopodobnie mają pochodzenie eukariotyczne, zostały odkryte w osadach jeziornych w południowej Afryce przez zespół Józefa Kaźmierczaka. Prezentują komórczakową budowę syfonalną. Datowane są one na ok. 2,8-2,7 mld lat[7].
Podział naturalny jądrowców przysparza wiele problemów z powodu niejasności w pokrewieństwie wielu grup systematycznych. Dawniejsze wnioskowanie o relacjach między poszczególnymi grupami na podstawie podobieństwa (głównie morfologicznego i anatomicznego) skutkowało wieloma błędami w związku z trudnością identyfikowania wpływu konwergencji i wtórnego upraszczania budowy. Od lat 80. XX wieku zaznaczył się wyraźny postęp w systematyce jądrowców. Wynika on z uwzględnienia takich źródeł o pokrewieństwie jak podobieństwo rRNA, grzebieni mitochondrialnych i aktyny. Mimo że wciąż wiele grup jądrowców oczekuje na ustalenie powiązań filogenetycznych, znane obecnie relacje pokrewieństwa pozwalają na wyróżnienie pięciu "supergrup" i określenie należących do nich linii rozwojowych[8][9][10]:
Dodatkowo Cavalier-Smith wyróżnia 2 taksony bez rangi[11]:
które są ponad taksonomicznie ponad supergrupami.
Podobne podejście przyjęto w pracy Adla i in. z 2019, gdzie eukarionty dzieli się na dwie główne grupy: Amorphea i Diaphoretickes oraz kilka mniejszych grup, których nie da się jednoznacznie przyporządkować do którejś z nich. Część z tych grup jest łączona w Excavata, przy czym w tym ujęciu jest to takson sztuczny, a jego status jako supergrupy jest podawany w wątpliwość[12].
System ten został w 2012 roku zastąpiony nowa poprawioną wersją, która uwzględniła zmiany jakie zaszły w tym czasie. System Adla z 2005 roku w następujący sposób systematyzował organizmy[13]:
Najbardziej popularny w XX w. podział jądrowców Roberta Whittakera i Lynn Margulis z 1978 r. wyróżniał 4 niższe taksony w randze królestw:
Niedoskonałość tego podziału związana jest z parafiletycznym charakterem grupy protistów.
Eukarionty, eukariota, eukarioty, organizmy eukariotyczne (Eukaryota, Eukarya), określane też jako jądrowce, jądrowe, organizmy jądrowe, karioty, kariota (Karyobionta) – organizmy zbudowane z komórek posiadających jądro komórkowe z chromosomami, co jest jednym z elementów odróżniających je od prokariontów. Jądro komórkowe odgraniczone jest od cytoplazmy podwójną błoną białkowo-lipidową. Nazwa naukowa pochodzi od greckich słów ευ ("dobry", "prawdziwy") i κάρυον ("orzech", "jądro").
W zależności od ujęcia systematycznego jądrowce są traktowane jako domena (cesarstwo, nadkrólestwo) lub królestwo. Najpopularniejszą z tych kategorii systematycznych jest obecnie domena. Do eukariotów zalicza się wszystkie organizmy komórkowe, z wyjątkiem bakterii i archeowców, a więc prokariotów. Szacunki dotyczące liczby gatunków w obrębie tej grupy wahają się pomiędzy 5 a 10 mln. Według analiz przeprowadzonych przez naukowców z Census of Marine Life, opublikowanych w sierpniu 2011 roku, liczba ta wynosi 8,7 (±1,3) mln, z czego naukowo opisane zostały mniej niż dwa miliony.
Komórki eukariotyczne są z zasady znacznie większe od prokariotycznych. Choć zmienność rozmiarów i jednych i drugich jest bardzo duża (najmniejsze eukarioty mają rozmiar mniejszy niż 1 µm), typowe proporcje objętości są rzędu tysiąc do jednego.
O domínio taxonômico Eukariota, Eukaria, Eukarya, Eukaryota, também referido como eucariotas ou eucariontes (do grego ευ, translit.: eu, "bom, perfeito"; e κάρυον, translit.: karyon, noz ou amêndoa, núcleo[3][4][5]) inclui todos os seres vivos com células eucarióticas, ou seja, com um núcleo celular cercado por uma membrana (DNA compartimentado, consequentemente separado do citoplasma) e com várias organelas. No núcleo está contida a maior parte do material genético, o DNA, enquanto uma parte menor está contida nas mitocôndrias.[6] Seu DNA está associado a proteínas histónicas.
Os eucariotas são, portanto, os organismos vivos unicelulares ou pluricelulares constituídos por células dotadas de núcleo, distinguindo-se dos procariotas (grupo parafilético), cujas células são desprovidas de um núcleo bem diferenciado.[7]
Variam desde organismos unicelulares até gigantescos organismos multicelulares, nos quais as células se diferenciam e desempenham funções diversas, não sobrevivendo isoladamente. Os eucariotas compartilham uma mesma origem, e por isso são agrupados numa hierarquia taxonômica superior ao reino: o domínio ou império, dependendo de como cada autor encara a origem dos eucariotas.
Fazem parte desta categoria de seres vivos, chamada de "Império Eucariota" por Cavalier-Smith, os Reinos: Animalia, Plantae, Fungi, Protista e Chromista ou Stramenopilla.[8] As formas vivas que não fazem parte do domínio Eukariota são aquelas incluídas nos reinos Bacteria e Archaea (anteriormente denominadas arqueobactérias), ou seja, os seres vivos com células procarióticas. Também há os vírus, que são seres acelulados.
As células eucarióticas são geralmente muito maiores que os procariontes e são muito mais compartimentadas.[9] Eles têm uma grande variedade de membranas com núcleos cercados pelo envelope nuclear, retículo endoplasmático e complexo de Golgi, além de mecanismos de brotamento e fusão de vesículas, incluindo exocitose e endocitose. Estruturas internas chamadas organelas são responsáveis por executar funções especializadas dentro da célula. Presença de lisossomos, peroxissoma e mitocôndrias.
Todos os eucariotos também são caracterizados por um esqueleto interno ou endosqueleto, neste caso chamado citoesqueleto,[10] composto de duas redes de proteínas: o sistema de microtúbulos e o sistema contrátil de actina/miosina, que desempenham um papel importante na definição da organização e forma celular, no tráfico intracelular (por exemplo, movimentos das vesículas e organelas) e na divisão celular.[11] O flagelo eucariótico característico e seus motores moleculares associados estão ancorados no citoesqueleto.
O DNA das células eucarióticas está contido em um núcleo celular separado do resto da célula por uma membrana dupla permeável. O material genético é dividido em vários blocos lineares chamados cromossomos, que são separados por um eixo microtubular durante a divisão celular. Os cromossomos contêm histonas, vários replicons, centrômeros e telômeros. Existe um ciclo celular característico com segregação mitótica e reprodução sexual por meiose. Inclui um complexo de poros nucleares, transporte transmembranal de RNA e proteínas através do envelope nuclear, íntrons e novos padrões de processamento de RNA usando o baço.
A origem dos eucariotas é uma das grandes questões atuais sobre Evolução e, até à data, carece de uma resposta definitiva. De modo semelhante à questão da origem da vida, nunca nos será possível determinar qual foi a sequência de eventos exata que levou à origem dos eucariotas, mas é-nos possível, de acordo com a melhor data científica existente, explorar quais as hipóteses mais prováveis.
Supõ-se que a origem dos eucariotas se deu uma única vez durante a história da vida na Terra.[12] Para alguns autores a origem da mitocôndria encontra-se diretamente associada à origem dos eucariotas.[13] Outros autores sugerem que cedo na história da Evolução os organismos se dividiram nos três domínios da vida (eucariotas, bactérias e arqueia), tendo adquirido a mitocôndria mais tarde. Estas duas hipóteses são contraditórias, e atualmente a maior parte da evidência suporta que a origem da mitocôndria se deu muito cedo na origem dos eucariotas, apoiando a hipótese de que o desenvolvimento deste organelo foi fundamental para o desenvolvimento dos eucariotas.[14]
Um dos argumentos para que a mitocôndria esteja na base da origem dos eucariotas é que todos os eucariotas estudados até ao momento têm, ou têm evidência de ter tido, mitocôndria.[15] Outros eucariotas possuem um organelo chamado de “hidrogenossoma”, que se encontra evolutivamente relacionado com a própria mitocôndria. O parasita Giardia é geralmente dado como exemplo de eucariota amitocondriado, sugerindo que se trata de um eucariota que surgiu cedo durante a evolução e que se separou dos restantes eucariotas antes da aquisição da mitocôndria. Estudos mostraram, no entanto, que a Giardia possui no seu genoma evidência de ter tido um organelo relacionado com a mitocôndria.[16] O facto de a Giardia não ter mitocôndria representa, portanto, não um caso de divergência antes da aquisição deste organelo, mas antes um caso de evolução redutiva (isto é, de perda de características – um fenómeno evolutivo comum em parasitas).
Uma das razões levantadas para sugerir que a aquisição da mitocôndria foi fundamental para o desenvolvimento da complexidade eucariótica é que apenas quando um organelo intracelular é utilizado para produzir energia é que é possível a um organismo aumentar de tamanho.[17] Isto encontra-se relacionado com o facto de que a energia produzia devido a processos quimiosmóticos (tais como a respiração celular) restringe o tamanho que os seres unicelulares podem adquirir. Em processos quimiosmóticos diversos processos de membrana ocorrem de modo a forçar a existência de um gradiente eletroquímico através de uma membrana celular. Por exemplo, uma cadeia respiratória pode utilizar a energia derivada da redução de compostos orgânicos (como açúcares) para transportar protões para o exterior da membrana. A diferença de concentração de protões entre o exterior e o interior da membrana causa uma tendência à entrada de protões novamente através da membrana. Os protões entram então através de proteínas associadas a ATP sintases, que usam assim a energia do gradiente para produzirem ATP – a moeda de troca energética comum a toda a vida. Este processo é análogo ao de uma central hidroelétrica, em que a queda de água causa a rotação de turbinas que resultam na produção de eletricidade. Todo este processos se encontra restrito pelo tamanho da membrana através da qual este processo possa ocorrer. Por este motivo, os organismos unicelulares só podem crescer até um determinado quociente entre a área membranar e o volume celular. Quando a energia passa a ser produzida em mitocôndrias no interior da célula esta restrição deixa de existir, passando os organismos a poder adquirir um maior tamanho e, consequentemente, maior complexidade.
Existem várias hipóteses que sugerem como é que a relação endossimbiótica que levou à origem dos eucariotas ocorreu. Uma destas hipóteses é chamada de Hipótese de Hidrogénio, proposta por Martin William e Miklós Müller em 1998.[18] Esta hipótese considera que a origem dos eucariotas se deve a uma relação endossimbiótica estabelecida entre uma arqueia (o hospedeiro) produtora de metano e uma bactéria produtora de hidrogénio. Como os metanogénios são organismos anaeróbicos que requerem hidrogénio, a relação endossimbiótica ter-se-á iniciado como uma relação sintrófica. Esta hipótese apresenta alguns problemas, tal como a dificuldade em explicar a razão pela qual a membrana plasmática eucariótica é mais semelhante à membrana plasmática bacteriana do que à membrana plasmática arqueana. Para além do mais, não explica a origem dos sistemas endomembranares eucarióticos nem a origem do núcleo.
A descoberta recente do filo arqueano Lokiarchaeota e de outros filos de arqueia (coletivamente chamados de superfilo Asgard) trouxe mais informações relativas à origem dos eucariotas.[19][20] Os organismos pertencentes a este superfilo são particularmente ricos em proteínas consideradas tipicamente eucariotas. Alguns autores sugerem que a existência destes genes indica que algumas espécies de arqueia já teriam complexidade quasi-eucariota, sugerindo que estes organismos representam os organismos mais próximos do hospedeiro original da endossimbiose que precedeu a origem dos eucariotas, e que estes organismos já teriam alguma da complexidade eucariótica, o que significa que esta hipótese representa uma hipótese intermédia entre aquela considera a possibilidade de existirem organismos amitocondriados, e a hipótese que considera que a mitocôndria é uma condição sine qua non para se considerar um organismo como eucariota. A descoberta do Superfilo Asgard levou ainda à sugestão de que os eucariotas e as arqueias formam um grupo monofilético, e consequentemente à existência de dois domínios apenas: as bactérias e um domínio que inclui as arqueias e os eucariotas. Esta hipótese é no entanto controversa e não existe ainda consenso na comunidade científica.[21][22]
Na antiguidade, as duas linhagens de animais e plantas foram reconhecidas. Eles receberam a classificação taxonômica do reino por Lineu. Embora ele tenha incluído os fungos nas plantas com algumas reservas, percebeu-se mais tarde que eles são bastante distintos e justificam um reino separado, cuja composição não era totalmente clara até a década de 1980.[23] Os vários eucariotos de célula única foram originalmente colocados com plantas ou animais quando se tornaram conhecidos. Em 1818, o biólogo alemão Georg A. Goldfuss cunhou a palavra protozoários para se referir a organismos como os ciliados,[24] e esse grupo foi expandido até abranger todos os eucariotos unicelulares e receber seu próprio reino, o Protista, por Ernst Haeckel em 1866.[25][26] Os eucariotos passaram a ser compostos de quatro reinos:
Os protistas eram entendidos como "formas primitivas" e portanto, um grau evolutivo, unidos por sua natureza unicelular primitiva. O desembaraço das divisões profundas na árvore da vida só começou realmente com sequenciamento de DNA, levando a um sistema de domínios ao invés de reinos, como o nível mais alto apresentado por Carl Woese, unindo todos os reinos eucariotos sob o domínio eucariótico.[27] Ao mesmo tempo, o trabalho na árvore protista se intensificou e ainda continua ativamente hoje. Várias classificações alternativas foram encaminhadas, embora não haja consenso no campo.
Eukaryotes são um clado geralmente considerado irmão de Heimdallarchaeota no grupo Asgard no Archaea. Os agrupamentos basais são o Opimoda, Diphoda, o Discoba e o Loukozoa. A raiz eucariota é geralmente avaliada como estando próxima ou mesmo em Discoba.
As árvores de ARNr construídas durante as décadas de 1980 e 1990 deixaram a maioria dos eucariotos em um grupo "coroa" não resolvido (não tecnicamente uma coroa verdadeira), que geralmente era dividida pela forma da crista mitocondrial. Os poucos grupos que carecem de mitocôndrias se ramificaram separadamente e portanto, acreditava-se que a ausência era primitiva; mas agora é considerado um artefato de atração de ramificações longas, e sabe-se que eles os perderam secundariamente.[28][29]
A partir de 2011, existe um amplo consenso de que os Rhizaria pertencem aos Stramenopiles e aos Alveolata, em um clado apelidado de supergrupo SAR, de modo que Rhizaria não é um dos principais grupos de eucariotos; também que os Amoebozoa e Opisthokonta são monofiléticos e formam um clado, frequentemente chamados de Unikonta.[30][31][32][33][34] Além disso, não parece haver consenso.
Foi estimado que pode haver 75 linhagens distintas de eucariotos. A maioria dessas linhagens são protistas.[35]
Os tamanhos conhecidos do genoma do eucariote variam de 8,2 megabases (Mb) em Babesia bovis a 112 000–220 050 Mb no dinoflagelado Prorocentrum micans, mostrando que o genoma do eucarioto ancestral sofreu uma variação considerável durante sua evolução. Acredita-se que o último ancestral comum de todos os eucariotos tenha sido um protista fagotrófico com um núcleo, pelo menos um centríolo e cílio, mitocôndrias facultativamente aeróbias, sexo (meiose e singamia), um cisto dormente com uma parede celular de quitina e/ou celulose e peroxissomos. A endossimbiose posterior levou à disseminação de plastídeos em algumas linhagens.
A seguinte árvore filogenética mostra as relações entre os principais grupos de Eukaryota segundo as propostas mais recentes de Cavalier-Smith:[36][37][38][39][40][41]
Eukaryota DiphodaProtista
Animalia
Fungi
Plantae
Chromista
O domínio taxonômico Eukariota, Eukaria, Eukarya, Eukaryota, também referido como eucariotas ou eucariontes (do grego ευ, translit.: eu, "bom, perfeito"; e κάρυον, translit.: karyon, noz ou amêndoa, núcleo) inclui todos os seres vivos com células eucarióticas, ou seja, com um núcleo celular cercado por uma membrana (DNA compartimentado, consequentemente separado do citoplasma) e com várias organelas. No núcleo está contida a maior parte do material genético, o DNA, enquanto uma parte menor está contida nas mitocôndrias. Seu DNA está associado a proteínas histónicas.
Os eucariotas são, portanto, os organismos vivos unicelulares ou pluricelulares constituídos por células dotadas de núcleo, distinguindo-se dos procariotas (grupo parafilético), cujas células são desprovidas de um núcleo bem diferenciado.
Variam desde organismos unicelulares até gigantescos organismos multicelulares, nos quais as células se diferenciam e desempenham funções diversas, não sobrevivendo isoladamente. Os eucariotas compartilham uma mesma origem, e por isso são agrupados numa hierarquia taxonômica superior ao reino: o domínio ou império, dependendo de como cada autor encara a origem dos eucariotas.
Fazem parte desta categoria de seres vivos, chamada de "Império Eucariota" por Cavalier-Smith, os Reinos: Animalia, Plantae, Fungi, Protista e Chromista ou Stramenopilla. As formas vivas que não fazem parte do domínio Eukariota são aquelas incluídas nos reinos Bacteria e Archaea (anteriormente denominadas arqueobactérias), ou seja, os seres vivos com células procarióticas. Também há os vírus, que são seres acelulados.
Protozoarele, ciupercile, plantele și animalele, sunt organisme eucariote. Acestea au celule cu structură complexă, în care materialul genetic este localizat în nucleul sau nucleii celulelor. În sistemele de clasificare noi (2015 - 2017) cu 7 regnuri și 3 domenii, regnurile eucariote sunt: Chromista, Fungi, Protista, Plantae, Animale. Dintre eucariote, organisme unicelulare sunt unele ciuperci și protozoarele, iar pluricelulare sunt în totalitate animalele și plantele. Celulele eucariotelor conțin organite citoplasmatice, așa cum sunt mitocondriile, cloroplastele etc.
Spre deosebire de eucariote, procariotele, regnul Monera și regnul Archaea nu au organite celulare protejate de membrane, așa cum ar fi aparatul Golgi și alte structuri complexe intracelulare.
Eukarya BikontaSubdiviziuni ale organismelor eucariote
Eukaryoty

Obazoa
CRuMs
Sar
Haptista
Vedecká klasifikácia prevažne podľa tohto článkuEukaryoty [1] (iné názvy: eukaryonty [2], eukaryontné/eukaryotické organizmy [2][3], jadrové organizmy[4], jadrovce[5], nukleobionty [6]; lat. Eukaryota/Eucaryota, Eukaryonta, Eukaryotae/Eucaryotae, Eukarya/Eucarya, Karyonta/Caryonta, Eucytota [7], Karyobionta [8], Nucleobionta [9]) sú organizmy, ktorých bunky (tzv. eukaryotické bunky) majú hlavnú genetickú informáciu uloženú v bunkovom jadre izolovanom od okolitého prostredia (cytoplazmy) fosfolipidovou membránou. Zahŕňajú všetky mnohobunkové organizmy, ale väčšina ich diverzity je tvorená drobnými jednobunkovými oraganizmy. Eukaryoty sú jednou z domén živých organizmov.
Bunky eukaryotov obsahujú menšie štruktúry, organely, ktoré sú tiež obalené fosfolipidovými membránami, preto ich nazývame organely endomembránovej sústavy. Okrem jadra sú to napríklad mitochondrie, plastidy a ďalšie. Toto je jeden z hlavných znakov, ktorým sa odlišujú od organizmov s jadrom neobaleným membránou, tzv. prokaryotov. Ďalší rozdiel medzi prokaryotmi a eukaryotmi je v ich ribozómoch (malých organelách slúžiacich na syntézu bielkovín): Cytoplazmatické ribozómy eukaryotov sú väčšie a ťažšie ako ribozómy prokaryotov. Ribozómy eukaryotov v plastidoch a mitochondriách sú však veľkosťou a hmotnosťou podobné ribozómom prokaryotov. Táto skutočnosť je jedným z dôkazov podporujúcich endosymbiotickú teóriu vzniku eukaryotických organizmov. Ďalší zásadný odlišný znak prokaryotov a eukaryotov je, že dedičná látka eukaryotov (DNA) je naviazaná na bázické bielkoviny.
Podľa často uvádzanej endosymbiotickej teórie vznikli eukaryotické bunky evolučne z prokaryotických buniek. Eukaryoty dosiahli od svojho vzniku pred 1,8 až 1,3 miliardami rokov značné množstvo foriem a patria medzi ne najväčšie organizmy na Zemi. Z taxonomického hľadiska ide spravidla o jednu zo základných skupín živých organizmov. Tradične sa členili na tieto základné taxóny: prvoky, chromista, rastliny, huby a živočíchy.[10] Aktuálne sa však užíva fylogenetická klasifikácia podložená molekulárnymi dátami. Podľa nej eukaryoty zaraďujeme do tzv. superskupín.
Bunky eukaryotov majú priemerne desaťkrát väčšie (lineárne) rozmery než bunky prokaryotických organizmov.[11] Väčšina eukaryotických buniek vrátane napr. telesných buniek človeka je však stále príliš malá na to, aby sa dala pozorovať voľným okom. Najmenšie eukaryotické bunky má zelená riasa Ostreococcus tauri, a to približne jeden mikrometer,[12] tzn. sú menšie ako napr. bunka baktérie Escherichia coli.[13] Iné eukaryotické bunky, napríklad niektoré mnohojadrové bunky vo vnútri tiel veľkých živočíchov (ako napr. niektoré nervové bunky), dosahujú zase obrovské rozmery, sú však neschopné samostatnej existencie. Pokiaľ sa však zameriame na najväčšiu bunku schopnú samostatnej existencie, existujú aj mimoriadne veľké jednobunkové organizmy. Veľmi veľká (až jeden meter) je jednobunková zelená riasa rodu Caulerpa.[14] Mnohojadrové plazmódiá prvokov, ako je Physarum polycephalum, môžu tiež dosahovať veľkosť niekoľko metrov – boli zaznamenaní jedinci vo vnútri tiel veľkých živočíchov s plochou 5,54 m².[15]
Bunky eukaryotov sú veľmi odlišné od prokaryotickej bunky nielen v otázke veľkosti, ale vykazuje značné rozdiely aj po štruktúrnej stránke. Na rozdiel od prokaryotov sú komplexnejšie a tzv. kompartmentalizované, teda rozdelené na jednotlivé kompartmenty („oddelenia“). Obsahujú napríklad pravé jadro obklopené dvojitou membránou, ktorá oddeľuje DNA od okolitých častí bunky. Ďalším výrazným znakom je endomembránový systém, čiže skupina organel taktiež obalených membránou. K týmto patrí hlavne endoplazmatické retikulum, Golgiho aparát, lyzozómy či mitochondrie, prípadne funkčné deriváty mitochondrií (hydrogenozómy, mitozómy). Obsahujú tiež tzv. cytoskelet, čo je akási vnútorná kostra bunky zaisťujúca pohyb buniek a vnútorný transport; plní tiež dôležité funkcie pri delení buniek. Nemenej dôležitou súčasťou sú ribozómy, u eukaryotov tzv. 80S (toto číslo udáva čas, za ktorý prebehne sedimentácia ribozomálnej makromolekuly pri jej ultracentrifugácii).
Okrem vyššie uvedených štruktúr, ktoré sú prítomné takmer vo všetkých eukaryotických bunkách, sa v mnohých skupinách eukaryotov vyvinuli aj iné špecifické organely, napríklad plastidy u rastlín a mnohých protíst. Na rozdiel od mitochondrií sa plastidy vyvinuli u niekoľkých, často nepríbuzných skupín eukaryotov. Tzv. primárne plastidy pochádzajú zo siníc a vznikli pravdepodobne len raz, a to u rastlín, ktoré v širšom poňatí zahŕňajú nielen zelené rastliny (Viridiplantae), ale aj červené riasy (Rhodophyta), glaukofyty (Glaucophyta) a tiež skupinu Rhodelphidia, ktorej zástupcovia sa však živí heterotrofne.[16] Plastidy u iných druhov fotosyntetizujúcich eukaryotov vznikli predovšetkým sekundárnou endosymbiózou, teda pohltením jednej z pôvodných skupín rastlín.
Obzvlášť u rôznych jednobunkových eukaryotov poznáme veľa ďalších unikátnych organel: vystreľovacie či vymršťovacie útvary (tzv. trichocysty), prichytávacie organely (rôzne stonky a prísavky), sťažiteľné, potravné a iné vakuoly, a podobne. Na pohyb zase slúžia rôzne bičíky a brvy (pozri kapitola pohyb). Úplne samostatnou kapitolou je anatómia mnohobunkových organizmov (pozri anatómia rastlín, anatómia živočíchov, anatómia húb).
Metabolické pochody výrazne ovplyvňuje už väčšia veľkosť eukaryotickej bunky, ktorá značne zvyšuje pomer objemu a plochy bunky.[17] V podstate rozoznávame u eukaryotov dva druhy výživy: obligátnu fotoautotrofiu u rastlín a iných fotosyntetizujúcich organizmov a obligátnu chemoheterotrofiu u živočíchov, húb a iných organizmov živiacich sa organickým materiálom.[18] Organizmus, ktorý dokáže využívať oba tieto základné typy výživy (napr. mnohé červenoočká), sa označuje termínom mixotrofný. Nájdeme však aj drobné odchýlky od klasických spôsobov získavania živín a energie. Napríklad niektoré prvoky a huby dokážu metabolizovať aj bez prístupu kyslíka.[19]
V rozmnožovaní (reprodukcii) eukaryotov hrá veľkú úlohu nepohlavné rozmnožovanie, ale evolučne unikátna je zrejme schopnosť rozmnožovať sa pohlavne (sexuálne). Pomerne často dochádza k pravidelnému striedaniu pohlavného a nepohlavného rozmnožovania, čiže k tzv. rodozmene (metagenézia). Len u niektorých protíst, ako Choanoflagellata, Euglenoidea, Trichomonadida a väčšina Arcellinida, nebola sexuálna reprodukcia doteraz pozorovaná a o jej existencii môžeme len špekulovať.[15]
Nepohlavne sa množí väčšina prvokov, ale aj niektoré mnohobunkové organizmy. U jednobunkových organizmov je nepohlavné rozmnožovanie spojené s mitózou (bunkovým delením). Pred vlastným rozdelením bunky (cytokinézou) musí byť DNA zmnožená a taktiež sa musia zmnožiť organely, aby každá dcérska bunka bola kompletná.
Rozlišuje sa niekoľko typov nepohlavného rozmnožovania, a to jednoduché delenie materskej bunky na dve dcérske (u protíst najbežnejšie), ďalej pučanie (napr. Suctoria) či mnohonásobné delenie bunky za vzniku množstva dcérskych buniek (rôzne Apicomplexa).[15] U mnohobunkových organizmov sa vyskytuje široké spektrum rôznych druhov nepohlavného rozmnožovania, napr. fragmentácia (u mnohých rastlín), tvorba spór nepohlavnou cestou, partenogenéza, apomixia alebo tiež pučanie.
Aj pohlavné rozmnožovanie je v rámci eukaryotickej domény (až na vyššie zmienené výnimky) neobyčajne rozšírené. V užšom zmysle pojmu pohlavné rozmnožovanie sú tohto typu rozmnožovania schopné spomedzi všetkých foriem života len eukaryoty. Typický scenár u mnohých prvokov a mnohobunkových organizmov vyzerá takto: v procese redukčného delenia (meiózy) vznikajú dva typy sexuálne diferencovaných buniek, tzv. gaméty, obvykle označované ako + a − (či samčie a samičie). Pokiaľ tieto bunky vyzerajú veľmi podobne, tak je rozmnožovanie izogamické, pokiaľ sú odlišné, hovoríme o anizogamii. Špecifickým typom je oogamia, pri ktorej je veľká samičia pohlavná bunka oplodnená malou pohyblivou samičou pohlavnou bunkou. Niekedy splývajú hneď rodičovské bunky (gamónty), príkladom je konjugácia nálevníkov.[15] Splynutím dvoch pohlavných buniek vzniká tzv. zygota, ktorá sa potom ďalej mitoticky delí. U niektorých mnohobunkových organizmov sa vyvinuli špecializované pohlavné orgány.
Charakter pohybu eukaryotických organizmov sa do značnej miery odvíja od ich telesnej stavby. Iný typ pohybu nájdeme u jednobunkových eukaryotov, inak sa samozrejme pohybujú mnohobunkovce.
U jednobunkových organizmov či jednobunkových štádií mnohobunkových organizmov (napr. spermie) hrajú hlavnú úlohu hlavne riasinky či bičíky, štruktúry s priemerom asi 0,2 mikrometra. Vlastné úzke telo (axonéma) je celé pokryté plazmatickou membránou a nachádza sa teda vo vnútri bunky. Na priečnom priereze je vidieť unikátne usporiadanie mikrotubúl (9×2+2), pri ktorom je jedna centrálna dvojica mikrotubulov obklopená kruhovite deviatimi ďalšími. Táto štruktúra je veľmi stabilná a líši sa len u niekoľkých skupín. Pohyb bičíkov a riasinek spotrebováva ATP a je založený na činnosti tzv. ATPáz. Poznáme však aj veľa iných štruktúr umožňujúcich pohyb samostatných buniek. Améboidný (meňavkovitý) pohyb eukaryotov je založený na polymerácii aktínu a vyskytuje sa napríklad u zástupcov ríše Amoebozoa či taktiež mnohých Foraminifera. Charakteristický druh pohybu u červenoočiek (Euglenida) zahŕňajúci vlnivé zmeny tela sa nazýva euglenoidný pohyb. Kĺzavým pohybom, ktorý je zrejme založený na cielenom vylučovaní slizu, sa môžu pohybovať napríklad Gregarinidea či Coccidea.[15] U mnohobunkových organizmov sa rozvinuli úplne iné druhy pohybu, väčšinou založené na kontrakcii špeciálnych typov buniek. Toto zahŕňa plazenie, chôdzu, beh, plávanie a podobne.
Genetický materiál v podobe DNA majú eukaryoty uložený v jadre, poprípade v niekoľkých jadrách (u nálevníkov) a v semiautonónmych organelách, ako sú mitochondrie a plastidy (vzácne tiež inde, napr. v nukleomorfe, zvyšku jadra po endosymbiontovi). Táto DNA sa následne, podobne ako u ostatných domén organizmov, prepisuje do RNA, ktorá potom slúži buď na syntézu proteínov (mRNA), alebo má určitú katalytickú funkciu ako taká (rRNA, tRNA a podobne).
V jadre tvorí DNA a niektoré proteíny (napr. históny) tzv. chromatín, ktorého jednotlivé oddelené časti sa nazývajú chromozómy. Veľkosť jadrového genómu (počet báz) je veľmi rozmanitá. Vo všeobecnosti platí, že čím zložitejší je organizmus, tým viac DNA obsahuje. Existujú však početné výnimky. Bunky niektorých rastlín majú napríklad desaťkrát viac báz, ako bunky človeka, bunky obojživelníkov zase môžu mať viac DNA ako bunky cicavcov.[20]
Najmenší eukaryotický genóm, obsahujúci len 2,9 miliónov párov báz, má vnútrobunková parazitická mikrosporídia menom Encephalitozoon cuniculi,[21][22] a to preto, že u nej došlo k redukcii genómu v rámci jej parazitického spôsobu života. Naopak, najväčší genóm zo všetkých známych eukaryotov (670 miliárd párov báz) má Amoeba dubia.[23] Genóm človeka tvorí 3 300 000 000 párov báz.[20]
Oproti prokaryotickým organizmom, u eukaryotov väčšina ich DNA nekóduje žiadny gén (a teda ani proteín). U ľudí tvorí nekódujúca DNA asi 97 % genómu a jej funkcie nie sú v drvivej väčšine prípadov dodnes známe. Označuje sa tiež ako junk DNA, súvisiace termíny sú tiež intróny, či repetitívna DNA.[17]
Mitochondriálna DNA a plastidová DNA sú v porovnaní s jadrovou DNA relatívne malé a uzavreté do kruhu. Považujú sa za pozostatok po endosymbiotickej udalosti, pri ktorej budúca eukaryotická bunka pohltila prokaryotický organizmus. Mitochondriálna DNA má veľa spoločných znakov s DNA alfa proteobaktérií, plastidová vykazuje príbuznosť s DNA cyanobaktérií (siníc).
Hoci vznik eukaryotických buniek je významný míľnik v evolučnej histórii života, čas jeho vzniku je stále neistý. Niekedy sa datuje do doby pred 1,8 – 1,3 miliardami rokov,[24] teda dávno pred kambrijskou a ediakarskou diverzifikáciou života. Inokedy sa dokonca uvádza doba pred dvoma miliardami rokov.[26] Z týchto období našej geologickej histórie je k dispozícii len málo fosílnych nálezov eukaryotov, ale existujú výnimky: boli popísané starohorné fosílne nálezy prvokov, ktoré sa označujú ako častí zástupcovia eukaryotov.[24] Taktiež niektoré nálezy rias (Grypania) sú podľa ich objaviteľov až 2,1 miliardy rokov staré.[25] Stopy po existencii eukaryotov sa dokonca našli aj v austrálskych bridliciach starých 2,7 miliardy rokov.[27] Napriek týmto nálezom sa hlavné súčasné línie eukaryotov pravdepodobne oddelili až ku koncu prekambria, konkrétne v období medzi 1,3 – 0,72 miliardami rokov. Fosílie raných eukaryotov, ako je napríklad Shuiyousphaeridium macroreticulatum, môže byť ťažké odlíšiť od prokaryotických buniek. U tohto pravdepodobného eukaryonta sa pri jeho zaraďovaní napríklad vychádzalo z veľkých rozmerov bunky, pevnej steny a výrastkov na jeho povrchu.[24] Väčšinu nálezov z proterozoika však nie sú vedci schopní zaradiť do konkrétnejšej skupiny eukaryotov. Kvalitné fosílie eukaryotov sa začínajú objavovať až v ediakare a kambriu, teda v období asi pred asi 700 – 600 miliónmi rokov.[28] V ediakare sa našli okrem zelených rias taktiež zvláštne planktónne organizmy zo skupiny Acritarcha.[29]
Pôvod eukaryotickej bunky doteraz nie je známy. Podľa súčasných poznatkov sa zdá, že ku vzniku eukaryotov prispeli tak archeálne, ako aj bakteriálne genómy. Jedným zo svetlých miest je vznik mitochondrií. V tejto organele bola totiž nájdená DNA, ktorá vykazuje značnú príbuznosť s baktériami z radu Rickettsiales. Endosymbiotická teória tak oprávnene tvrdí, že mitochondrie vznikli práve pohltením Rickettsií inou bunkou a dnešné mitochondrie sú potomkovia týchto baktérií,[30] ktoré sa ale dlhodobým vývojom vo vnútri eukaryotických buniek značne zmenili.
Samotný vznik mitochondrií však neznamenal vznik eukaryotov v dnešnom slova zmysle. Eukaryogenézu sa snaží ďalej vysvetliť viac než 20 hypotéz.[31] Podľa archeálnej hypotézy vznikla protoeukaryotická bunka z bunky istej archebaktérie a mitochondrie sa vyvinuli až za určitý čas, a to pohltením bakteriálneho symbionta.[30] Detailnejšie sa okolnosťami archeálnej hypotézy zaoberá tzv. vodíková hypotéza, podľa ktorej spomínaná archebaktéria metabolizovala vodík, ktorý jej práve poskytovali v rámci svojho bunkového dýchania endosymbiotické baktérie. Tieto baktérie následne prešli evolúciou a zmenili sa na mitochondrie.[32]
Niektoré štúdie však navrhujú o niečo zložitejší spôsob vzniku eukaryotickej bunky. Takzvané chimérické (čiže fúzne) hypotézy predpokladajú, že amitochondriálny eukaryont (čiže predok eukaryotov ešte bez mitochondrií) vznikol splynutím jednej bunky archeálnej a jednej bakteriálnej. Eukaryotický genóm je podľa tejto hypotézy tiež mixom archeálnych a bakteriálnych génov. Až potom vznikli mitochondrie (a plastidy), a to ďalšími endosymbiotickými udalosťami.[33] Podobná teória navrhuje iný sled udalostí: archebaktéria strávila baktériu ako potravu a časť bakteriálnej DNA sa včlenila do archeálnej DNA.[34] Úplne iný pohľad ponúka teória „Neomura“, ktorá považuje archea i eukaryota za potomkom určitej zložitej grampozitívnej baktérie, konkrétne istej aktinobaktérie. Táto aktinobaktéria prešla značným vývojom a až neskôr endosymbioticky získala mitochondrie.[35] Nedostatkom týchto teórií je fakt, že v súčasnosti nepoznáme žiadne eukaryotické organizmy primárne bez mitochondrií, hoci ich existenciu tieto teórie predpokladajú.[30]
U eukaryotov mnohobunkovosť vznikla niekoľkokrát nezávisle od seba, zvlášť u živočíchov, rastlín, húb a mnohých ďalších eukaryotických taxónov.[37] Vznik (respektíve vzniky) mnohobunkovosti bol značným evolučným úspechom eukaryotov. Vyriešená bola napríklad otázka vzájomného dorozumievania buniek a vzájomnej deľby práce.[38] U mnohobunkových živočíchov sú unikátne nielen gény zaisťujúce správny embryonálny vývoj (napr. hox gény), ale aj gény zaisťujúce komunikáciu medzi bunkami. U mnohobunkovcov sa vyskytujú aj nové bunkové štruktúry nepozorované u jednobunkovcov (dezmozómy a iné bunkové spoje).[39]
Najstaršie mnohobunkové živočíchy, ktoré sú dnes známe, žili v období ediakara niekedy pred 570 – 550 miliónmi rokov,[39] ako napovedajú aj niektoré údaje z molekulárnej biológie. Zreteľnejší fosílny záznam sa však objavuje až v kambriu. Z tohto obdobia, označovaného aj ako kambrická explózia druhov, pochádza množstvo fosílnych dokladov súčasných živočíšnych kmeňov.[40][41] Táto „explózia“, však môže byť iba zdanlivá, pretože podstatným rozdielom kambrických a prekambrických mnohobunkovcov, je objavenie sa pevných schránok, ktoré sa podstatne lepšie zachovávajú v usadených horninách. To malo za následok, že na rozhraní ediakary a kambria sa naraz objavujú bohaté, rozmanité fauny v lokalitách takmer na celom zemskom povrchu.[28] U rastlín (vrátane mnohých červených rias a zelených rias) sa mnohobunkovosť vyvinula hneď niekoľkokrát, a to zrejme v súvislosti s tým, že fotosyntetizujúce organizmy sa nemusia toľko pohybovať za potravou.[42] Vývoj mnohobunkového usporiadania sa u rastlín uberal troma základnými cestami. Niektoré kolónie zelených rias, aké tvorí napr. váľač gúľavý (Volvox globator), sa sformovali z niekoľkých pôvodných jedincov. Inokedy sa v pôvodnej bunke mnohonásobne delilo jadro, čím došlo k vzniku jednobunkových, ale viacjadrových stielok (napr. rod Caulerpa). Tretí spôsob vzniku mnohobunkových rastlín je ten, že pôvodná bunka sa mitoticky delila a jednotlivé dcérske bunky sa špecializovali na určité funkcie.[17] U húb nie je možné pozorovať nejaký všeobecný trend k mnohobunkovosti a nie je úplne jasné, koľkokrát v rámci húb mnohobunkovosť vznikla.[39]
Taxónov, u ktorých existuje mnohobunkovosť, je však oveľa viac. Príkladom sú akrázie (Acrasidae), chaluhy (Phaeophyceae), vajcové huby (Oomycota), pravé slizovky (Eumycetozoa) a dokonca niekoľko nálevníkov (Ciliophora) tvoriacich plodničky.[37][42]
Eukaryotické organizmy žijú takmer všade, kde je dostatočný konštantný prísun energie, a to buď slnečného žiarenia pre autotrofov, alebo dostatok živín pre výživu heterotrofov. Extrémofilné eukaryoty, podobne ako mnohé baktérie a archea, žijú aj na mnohých neobvyklých stanovištiach s extrémnymi podmienkami. Obmedzujúcim faktorom je pre eukaryotické organizmy hlavne teplota prostredia, ktorá na rozdiel od baktérií a archeí (ani u väčšiny termofilných eukaryotov) nesmie presiahnuť 70 °C.[43]
Zoznam extrémnych prostredí je pomerne dlhý. Poznáme anaeróbne eukaryoty (mnohé prvoky s hydrogenozómami, kvasinky), termofily (napr. červená riasa Cyanidium caldarium, termofilné huby z kompostov, niektoré mnohoštetinavce), chladnomilné eukaryoty (psychrofilné prvoky z arktických a antarktických morí), acidofily (Cyanidium caldarium a niekoľko húb žijúcich v pH 0), alkalofily (niektoré prvoky žijúce v pH 10), halofily (napr. riasa Dunaliella salina), barofily (hlbokomorské holotúrie v 110 MPa) a xerofily (mnohé púštne huby a lišajníky).[43]
Množstvo eukaryotických organizmov sa vyznačuje parazitickým spôsobom života, a to často bez ohľadu na ich vzájomnú nepríbuznosť. V centre pozornosti vedcov sú hlavne jednobunkové parazity človeka. Z ríše Excavata patrí medzi parazity mnoho diplomonád (Diplomonadida, napr. Giardia intestinalis) a trichomonád (Trichomonadida, napr. Trichomonas vaginalis), ďalej trypanozómy (Trypanosomatida, najmä rody Trypanosoma a Leishmania, pôvodcovia niektorých známych ľudských ochorení), či niektorí zástupcovia triedy Heterolobosea (napr. Naegleria fowleri). Z ríše Chromalveolata sú známe niektoré parazitické nálevníky (Ciliophora) a ďalej hlavne mnohé Apicomplexa, ako kokcídie (Coccidea, vrátane Toxoplazmy) a krvinkovky (Haematozoea, napr. rod Plasmodium, pôvodca malárie). Z ríše Rhizaria sa vyznačujú parazitickým spôsobom života napríklad nádorovky (Phytomyxea) či haplosporídie (Haplosporidia). Z ríše Amoebozoa medzi cudzopasníky patrí napríklad Acanthamoeba, Entamoeba histolytica a ďalšie. Poznáme i mnohé parazitické rastliny, konkrétne jednobunkové riasy rodu Prototheca, ale aj mnohé vyššie rastliny, ako Cuscuta či poloparazitické imelo (Viscum). Z vetvy smerujúcej k živočíchom a hubám (Opisthokonta) je tiež veľa známych parazitov: napríklad Mesomycetozoa a Myxozoa. Z ríše huby (Fungi) parazitujú mikrosporídie (Microspora), rôzne plesne (napr. rody Aspergillus, Penicillium) a kvasinky (napr. Candida albicans). Zo živočíchov (Metazoa) parazitujú mnohí zástupcovia kmeňa ploskavcov (Platyhelminthes), ako ploskule (Turbellaria), pásomnice (Cestoda), motolicovce (Monogenea) a motolice (Trematoda). Ďalej sa paraziti nachádzajú medzi kmeňmi háčikohlavce (Acanthocephala), obrúčkavce (Annelida, hlavne pijavice, Hirudinea) a tiež v rámci kmeňa hlístovce (Nematoda) a strunovce (Nematomorpha). Existujú však aj parazitické článkonožce (Arthropoda) a chordáty (Chordata).[26]
Okrem parazitizmu sa u eukaryotov prirodzene vyskytuje široké spektrum ďalších symbiotických vzťahov, ako je mutualizmus (obojstranne prospešný) alebo komenzalizmus (pre jedného symbionta prospešný, pre druhého neutrálny). Na jednej strane eukaryoty často poskytujú svoje telo pre rôznych endosymbioticky žijúcich eukaryotov či prokaryotov, konkrétnym príkladom sú prvoky vo vnútri tráviacej sústavy prežúvavcov. Nájdeme však aj mnohé ďalšie symbiotické vzťahy, ako sú početné symbiotické vzťahy mravcov, lišajníky (symbióza huby s riasou), mykoríza (niektoré vyššie rastliny s hubami), či spolupráca medzi mnohými kvitnúcimi rastlinami a ich opeľovačmi, ktorí prenášajú peľ z kvetu na kvet.[44]
Historické snahy o klasifikáciu živých organizmov sú z veľkej časti práve klasifikáciou eukaryotov, keďže len medzi eukaryotmi sa vyskytujú veľké organizmy, ktoré boli známe od počiatkov histórie ľudstva. Na najvyššej úrovni v priebehu času dochádzalo k veľkým zmenám. Zatiaľ čo Linné rozoznával len dve základné ríše, rastliny a živočíchy, postupne sa ukázalo, že tieto dve skupiny (a tiež niektoré jednobunkovce a huby) sú si v mnohých oblastiach vzájomne podobné a zároveň diametrálne odlišné od baktérií. Pravdepodobne prvým, kto si to uvedomil, bol francúzsky biológ Edouard Chatton. Ten v roku 1937 navrhol deliť všetky živé organizmy na dve ríše, eukaryota a prokaryota. Jeho návrh sa však neujal a znovu sa k nemu v roku 1962 vrátili Roger Stanier a Cornelius Van Niel. Hoci v súčasnosti je termín prokaryota považovaný za zastaraný a nerešpektujúci fylogenetický vývoj, taxón eukaryota sa bežne používa.[45] V roku 1990 ho podporili Carl Woese, Otto Kandler a Mark Wheelis, ktorí rozčlenili všetok život do troch domén, Archaea, Bacteria a Eucarya.[46]
Tradične sa eukaryoty delili na niekoľko ríší, napríklad podľa Whittakera[47] na ríše živočíchy (Animalia), rastliny (Plantae), huby (Fungi) a Protista. Cavalier-Smith ďalej od nich oddelili organizmy, ktoré získali plastid pohltením červené riasy, ako novú ríšu Chromista.[48] Dnes sa však presadzujú úplne odlišné systémy.[49]
LinnaeusDonedávna bolo bežné členenie organizmov do troch domén z roku 1990.[54][55] Eukaryota a Archea boli spoločne zaraďované do skupiny Neomura. Tieto dve domény sú si naozaj navzájom príbuzné, ale dnes sa väčšinou eukaryoty kladú ako súčasť taxónu ASGARD priamo uprostred archeí.[35][56][57]
Klasifikácia eukarotov na úroveň niekdajších ríší sa začína pomaly ustáľovať na systéme takzvaných superskupín. Zatiaľ čo napríklad väčšina učebníc sa stále drží taxónov ako prvoky alebo chromista,[54] aktuálne systémy s týmito termínmi takmer vôbec či vôbec nepracujú.[58] Do centra pozornosti sa stavia niekoľko taxónov označovaných ako „supergroups“,[59] ktoré zaraďujeme ešte ďalej do väčších nadradených taxónov. Aktuálny systém publikovaný Sinom M. Adlom pracuje k roku 2019 s deviatimi superskupinami:[60][61]
Nadradené skupiny Superskupina Zaraďované skupiny Popis Opimoda Amorphea Amoebozoa (meňavkovce) Evosea, Discosea, Tubulinea Zahŕňa väčšinu meňavkovitých organizmov a slizovky. Obazoa Opisthokonta, Breviatea, Apusomonadida Názov odkazuje na jej tri súčasti. Opistokonta sú typické zadným bičíkom a zahŕňajú známe mnohobunkové organizmy ako živočíchy či huby. CRuMs[62] Collodictyonidae, Rigifilida, Mantamonas Drobná skupina skupina jednobunkových organizmov, názov je slovnou hračkou. Diphoda[63] Diaphoretickes Sar Stramenopila, Alveolata, Rhizaria Skupina združuje jadrá niekdajších skupín Protista a Chromista vrátane mnohobunkových chalúh. Archaeplastida(rastliny)
Viridiplantae, Rhodophyta, Glaucofyta, Rhodephidia[64] Pravdepodobne monofyletický taxón obsahujúci mnohobunkové rastliny a ich jednobunkových príbuzných. Cryptista(kryptomonády)
Cryptomonadida, Katablepharida, Palpitomonas Kryptomonády v širšom slova zmysle. Haptista Haptophyta, Centrohelida Ide v podstate o haptophyta rozšírené o centrohelidné slnivky. Hemimastigophora[65] Drobná jednobunková skupina. Excavata(exkaváty, vykotlance[66])
Discoba, Metamonada Organizmy s cytosómom v podobe ryhy, ktorý je typický pre prvé eukaryoty[67]. Skupina nemusí byť monofyletická, Discoba a Metamonada nemusia byť sesterskými taxónmi.Aj napriek relatívne dobre zmapovaným príbuzenským vzťahom dnes mnoho skupín a rodov nedokážeme zaradiť do žiadnej superskupiny. Sina M. Adl a jeho kolektív takéto organizmy uvádzajú mimo publikovanú klasifikáciu ako Eukaryota incertae sedis, aby sa predčasne zabránilo vzniku polyfyletických taxónov.[68]
Tento článok je čiastočný alebo úplný preklad článku Eukaryota na českej Wikipédii.
Eukaryoty (iné názvy: eukaryonty , eukaryontné/eukaryotické organizmy , jadrové organizmy, jadrovce, nukleobionty ; lat. Eukaryota/Eucaryota, Eukaryonta, Eukaryotae/Eucaryotae, Eukarya/Eucarya, Karyonta/Caryonta, Eucytota , Karyobionta , Nucleobionta ) sú organizmy, ktorých bunky (tzv. eukaryotické bunky) majú hlavnú genetickú informáciu uloženú v bunkovom jadre izolovanom od okolitého prostredia (cytoplazmy) fosfolipidovou membránou. Zahŕňajú všetky mnohobunkové organizmy, ale väčšina ich diverzity je tvorená drobnými jednobunkovými oraganizmy. Eukaryoty sú jednou z domén živých organizmov.
Bunky eukaryotov obsahujú menšie štruktúry, organely, ktoré sú tiež obalené fosfolipidovými membránami, preto ich nazývame organely endomembránovej sústavy. Okrem jadra sú to napríklad mitochondrie, plastidy a ďalšie. Toto je jeden z hlavných znakov, ktorým sa odlišujú od organizmov s jadrom neobaleným membránou, tzv. prokaryotov. Ďalší rozdiel medzi prokaryotmi a eukaryotmi je v ich ribozómoch (malých organelách slúžiacich na syntézu bielkovín): Cytoplazmatické ribozómy eukaryotov sú väčšie a ťažšie ako ribozómy prokaryotov. Ribozómy eukaryotov v plastidoch a mitochondriách sú však veľkosťou a hmotnosťou podobné ribozómom prokaryotov. Táto skutočnosť je jedným z dôkazov podporujúcich endosymbiotickú teóriu vzniku eukaryotických organizmov. Ďalší zásadný odlišný znak prokaryotov a eukaryotov je, že dedičná látka eukaryotov (DNA) je naviazaná na bázické bielkoviny.
Podľa často uvádzanej endosymbiotickej teórie vznikli eukaryotické bunky evolučne z prokaryotických buniek. Eukaryoty dosiahli od svojho vzniku pred 1,8 až 1,3 miliardami rokov značné množstvo foriem a patria medzi ne najväčšie organizmy na Zemi. Z taxonomického hľadiska ide spravidla o jednu zo základných skupín živých organizmov. Tradične sa členili na tieto základné taxóny: prvoky, chromista, rastliny, huby a živočíchy. Aktuálne sa však užíva fylogenetická klasifikácia podložená molekulárnymi dátami. Podľa nej eukaryoty zaraďujeme do tzv. superskupín.
Evkarionti (tudi evkarioti ) (znanstveno ime Eukaryota) so organizmi, ki jih gradijo evkariontske (evkariotske) celice oziroma evcite.
Na Zemlji obstajata dve veliki skupini organizmov: prokarionti in evkarionti. Razlika med tema dvema vrstama življenja je zelo velika, čeprav oboji temeljijo na istih molekularno bioloških osnovah, natančneje: v obeh primerih je osnovna enota življenja celica, ki jo sestavlja ista skupina organskih makromolekul (proteini, sladkorji, lipidi in nukleinske kisline), ki je udeležena v istih osnovnih biokemičnih procesih. Najbolj očitna razlika med prokarionti in evkarionti je na nivoju celične organizacije in strukture, evkariontska celica je tako precej večja in bolj kompleksna od prokariontske. Poleg tega so lahko evkariontski organizmi večcelični, medtem ko so prokarionti samo v enocelični obliki. Evkarionti so praktično vse rastline in živali, medtem ko so prokarionti bakterije in arheje.
Obstajajo tri vrste evkariontske celice, živalska, rastlinska in celica glive, ki se nekoliko razlikujejo v celični zgradbi. Tipična evkariontska celica je ovalne oblike in velika 100 μm. Osnovna struktura, ki določa njeno velikost in obliko ter jo ločuje od okolice, je celična membrana. Rastlinska celica ima okrog membrane dodaten zaščitni sloj, t. i. celično steno iz celuloze. V notranjosti celice ali citosolu se nahajajo številni celični organeli. To so specializirane strukture obdane z lastno membrano, ki opravljajo pomembne celične funkcije. Večina organelov je skupna tako rastlinski kot živalski celici, rastlinske celice pa imajo dva dodatna organela, ki jih živalske nimajo, namreč kloroplaste in vakuole. Poleg organelov se v citoplazmi prosto nahajajo številni encimi in druge beljakovine, ki so udeleženi v celičnih procesih, kot sta sinteza in razgradnja beljakovin.
Evkarionti (tudi evkarioti ) (znanstveno ime Eukaryota) so organizmi, ki jih gradijo evkariontske (evkariotske) celice oziroma evcite.
Na Zemlji obstajata dve veliki skupini organizmov: prokarionti in evkarionti. Razlika med tema dvema vrstama življenja je zelo velika, čeprav oboji temeljijo na istih molekularno bioloških osnovah, natančneje: v obeh primerih je osnovna enota življenja celica, ki jo sestavlja ista skupina organskih makromolekul (proteini, sladkorji, lipidi in nukleinske kisline), ki je udeležena v istih osnovnih biokemičnih procesih. Najbolj očitna razlika med prokarionti in evkarionti je na nivoju celične organizacije in strukture, evkariontska celica je tako precej večja in bolj kompleksna od prokariontske. Poleg tega so lahko evkariontski organizmi večcelični, medtem ko so prokarionti samo v enocelični obliki. Evkarionti so praktično vse rastline in živali, medtem ko so prokarionti bakterije in arheje.
Eukaryoter (Eukaryota) utgör en av de tre domäner som organismer delas in i enligt modern systematik. En eukaryot är en organism som har en eller flera komplexa celler i vilka arvsmassan återfinns i en cellkärna avgränsad av ett cellmembran. Dessa celler är också relativt stora (10–100 mikrometer).
Eukaryoter omfattar traditionellt rikena djur, växter, svampar, samt protister, vilket i själva verket är en samlingskategori för eukaryoter som ej passar in i övriga riken. Eukaryoterna kontrasteras mot prokaryoterna (bakterier och arkéer), vilka saknar cellkärna och andra komplexa cellstrukturer.
Ordet är bildat av grekiskans εὖ (eu, ’god’, ’äkta’) och κάρυον (karyon, ’nöt’, ’kärna’) vilket syftar på cellkärnan.
De äldsta fossilen av eukaryoter är 1,8–1,9 miljarder år gamla och har hittats i Kina[2], och rimligtvis ligger eukaryoternas ursprung ytterligare något längre tillbaka i tiden, kanske så mycket som tre miljarder år sedan[3].
Det finns flera olika hypoteser om exakt hur den eukaryota cellen först utvecklades. Inslag av endosymbios, det vill säga att flera olika bakterier gick ihop till en gemensam cell, är ett framträdande inslag i de flesta hypoteser; se endosymbiontteorin. Vad gäller mitokondrier och kloroplaster är endosymbiosen välbelagd och det har i båda fallen gått att identifiera, vilken bakterie som inkorporerats (α-proteobakterier respektive cyanobakterier). Vad gäller cellkärnan är osäkerheten stor och ingen hypotes är allmänt accepterad. Ett endosymbiotiskt ursprung för cellkärnan utgår från en prokaryot, vars cellmembran veckas inåt till dess att det omslutit arvsmassan och bildat kärnmembran och endoplasmatiskt nätverk. Andra prokaryota bakterier med effektiv energiproduktion och cyanobakterier med fotosyntes tränger in i cellen och drar med sig lite av cellmembranet, vilket förklarar att mitokondrien och kloroplasten har dubbla membran. Mitokondrien, ribosomen och andra organeller började fungera tillsammans. Både mitokondrien och kloroplasten har egen arvsmassa och kopieras självständigt.
Eukaryoter utvecklade meios, sexuell reproduktion, för cirka 1,2 miljarder år sedan, och det var då evolutionstakten exploderade.[källa behövs]
Domänen eukaryoter (Eukaryota) har traditionellt indelats i fyra riken (med prokaryoterna i ett eget rike):
Denna indelning är dock föråldrad, och motsvarar inte de faktiska släktskapsförhållandena. Senare arbete, främst med dna-data, pekar snarare på en indelning i fem supergrupper även om alla detaljer mellan och inom grupperna inte är helt klarlagda:[4][5][1]
Opisthokonta djur, svampar, choanoflagellater etc. Amoebozoa de flesta amöbor och slemsvampar SAR Stramenopiles, Alevolata och Rhizaria Excavata diverse flagellater Archaeplastida växter, grönalger, rödalger och glaukofyterDärutöver har det föreslagits att eukaryoter kan klassificeras i två större grupper, Unikonta och Bikonta,[6] som härstammar från organismer med en respektive två flageller. I det här systemet är ophistokonter och amöbor (Amoebozoa) unikonter, och resten är bikonter.
Eukaryoter (Eukaryota) utgör en av de tre domäner som organismer delas in i enligt modern systematik. En eukaryot är en organism som har en eller flera komplexa celler i vilka arvsmassan återfinns i en cellkärna avgränsad av ett cellmembran. Dessa celler är också relativt stora (10–100 mikrometer).
Eukaryoter omfattar traditionellt rikena djur, växter, svampar, samt protister, vilket i själva verket är en samlingskategori för eukaryoter som ej passar in i övriga riken. Eukaryoterna kontrasteras mot prokaryoterna (bakterier och arkéer), vilka saknar cellkärna och andra komplexa cellstrukturer.
Ordet är bildat av grekiskans εὖ (eu, ’god’, ’äkta’) och κάρυον (karyon, ’nöt’, ’kärna’) vilket syftar på cellkärnan.
 Ökaryot hücrelerdeki temel çekirdek yapısının şeması. Bilimsel sınıflandırma Üst âlem: Eukaryota
Ökaryot hücrelerdeki temel çekirdek yapısının şeması. Bilimsel sınıflandırma Üst âlem: EukaryotaÖkaryotlar (Latince: Eukaryota), hücrelerinde bir çekirdek ve başka organeller içeren bir canlılar grubu olup bilimsel sınıflandırmada arkeler ve bakterilerle beraber tüm canlıları kapsayan üç ana gruptan biridir.
Ökaryotların genetik malzemeleri zarla çevrili çoğunlukla bir, nadiren birden çok çekirdek içinde yer alırlar. Bu nedenle kelime, gerçek (Grekçe: eu) ve çekirdek (Grekçe: karyon) sözcüklerinden türetilmiştir. Sıfat hâli ökaryotiktir. Bakteri ve arkeler çekirdeksiz olduklarından beraberce prokaryot olarak adlandırılırlar (evvel (Grekçe: pro-) ve çekirdek (Grekçe: karyon)). Çekirdeğin yanı sıra ökaryotların kloroplast veya mitokondri gibi zarla çevrili çeşitli organelleri vardır. Bu tür hücre içi karmaşık yapılar da prokaryotlarda bulunmaz.
Ökaryotların ortak bir atası olduğu için bir üst âlem (İngilizce: domain) olarak tanımlanmışlardır. Üst âlem sisteminde ökaryotların prokaryotlara kıyasla arkelerle daha çok ortak özellikleri olduğundan arkelerle beraber neomura kladı içinde gruplandırılırlar.
Ökaryotlar genel olarak bitki, hayvan, mantar ve Protista olarak dört gruba ayrılırlar. Ancak protista grubu aslında bitki, hayvan ve mantar olarak sınıflandırılamayan canlıları bir arada toplayan bir grup olduğu için bazı biyologlar tarafından kabul görmez ve yerine daha küçük gruplar tanımlarlar.
Çok çeşitli ökaryotik hücre tipi olmakla beraber hayvan ve bitkilerin en yaygın ve iyi bilinen çeşitleri olduklarından ökaryot yapısının anlaşılması için iyi bir başlangıç noktası oluştururlar. Ancak mantar ve çoğu protistanın hayvan ve bitkilerden önemli farklılıkları vardır.
Hayvan hücresi, hayvanların dokularını oluşturan bir ökaryotik hücre tipidir. Hayvan hücreleri diğer ökaryotlardan (özellikle bitki hücrelerinden) belirgin bir farklılık gösterir; hücre duvarı ve kloroplastları yoktur ve kofulları daha küçüktür. Hücre zarı esnek olduğundan hayvan hücreleri çeşitli şekillere girebilir ve fagositik bir hücre, başka cisimleri içine alabilir. Ayrıca bitki hücresinden farklı organeleri vardır. İnsan hücreleri biyolojik olarak ökaryotik hücrelerle aynı kategoriden sayılırlar.
Bitki hücreleri diğer ökaryotik organizmaların hücrelerinden oldukça farklıdırlar. Belirgin özellikleri
Mantar hücreleri en çok hayvan hücrelerine benzerler. Hayvan hücrelerinden farklılıkları
Ökaryotik hücreler, prokaryotlardan genelde çok daha büyüktürler. Organel olarak adlandırılan çeşitli iç zarlar ve iç yapılar, ayrıca miktrotüpçük (İngilizce: microtubule), mikroiplik (İngilizce: microfilament) ve ara ipliklerden (İngilizce: intermediate filaments) oluşmuş hücre iskeletine sahiptirler. Hücre iskeleti, hücrenin iç yapısını ve şeklini belirler. Ökaryotik DNA, hücrenin bölünme dönemlerinde kromozom olarak adlandırılan doğrusal tomarlar hâline dönüşür. Bunlar çekirdeğin bölünmesi sırasında kopyalanıp mikrotüpçükler tarafından çekilirler. Eşeysiz hücre bölünmesine (mitoz) ek olarak çoğu hayvan hücresinin hücre kaynaşması yoluyla gerçekleşen bir eşeyli üreme sürecine (mayoz) sahiptir. Bu üreme süreci prokaryotlarda görülmez.
Ökaryotik hücrelerde çeşitli zarla çevrili yapılar bulunur. Bunlara toplu olarak iç zar sistemi denir. Vezikül veya koful (vaküol) gibi basit bölmeler başka zarlardan tomurcuklanarak meydana gelir. Çoğu hücreler endositoz adı verilen bir süreçle besin ve diğer maddeleri içlerine alırlar; endositozda dış zar içe kıvrılıp sonra büzülerek bir vezikül oluşturur. Çoğu zarla çevrili organelin evrim sırasında bu tür veziküllerden meydana gelmiş olduğu varsayılmaktadır.
Çekirdek, çekirdek kılıfı olarak adlandırılan bir çift zar ile çevrilidir. Çekirdek örtüsünde bulunan gözenekler moleküllerin girip çıkmasını sağlar. Çekirdek kılıfının çeşitli tüp veya yapraksı uzantıları, endoplazmik retikulum (ER) olarak adlandırılan yapıyı oluşturur. Bu yapı protein ulaşımı ve olgunlaşmasından sorumludur. Granüllü ER'da ribozomlar bulunur ve bunların sentezlediği proteinler ER'un iç kısmına (lümenine) girer. Bu proteinler sonradan düz ER'dan (yani ER'un granülsüz kısmından) tomurcuklanan veziküllerin içine girerler. Çoğu ökaryotta bu protein taşıyan veziküller golgi aygıtı veya diktiyozom denen yassılaşmış vezikül desteleriyle kaynaşırlar ve proteinler orada çeşitli yapısal değişimlere uğrarlar.
Veziküller çeşitli amaçlar için özelleşmiştir. Örneğin lisozomlar, besin kofullarının içindekileri sindiren enzimler bulundurur, peroksizomlarsa hücre için zehirli olan peroksiti parçalar. Çoğu protozoada bulunan büzülür (kontraktil) kofullar, hücredeki fazla suyu toplayıp dışarı atarken ekstruzomlar avcı (predatör) canlıları kaçırmasına ve hücrenin avını yakalaması için dışarı madde atmasına yarar. Çok hücreli canlılarda hormonlar, çoğu zaman veziküllerde üretilirler. Yüksek bitkilerde hücre hacminin büyük bir kısmını işgal eder merkezî koful, hücrenin osmotik basıncını sabit tutar.
Mitokondriler hemen her ökaryotta bulunan organellerdir. Bir çift zarla çevrilidirler ve içte olanına krista adı verilen iç kıvrımlardan oluşurlar. Hücre solunumu burada gerçekleşir. Kendi DNA'ları vardır ve başka mitokondrilerin bölünmesi sonucu meydana gelirler. Evrim sırasında endosimbiotik bakterilerden (proteobakteriler) oluştukları teorisi genel kabul görmüştür. Mitokondrisi olmayan birkaç protozoda da hidrojenozom ve mitozomlar gibi mitokondri türevi organeller mevcuttur.
Bitki ve çeşitli alg gruplarında ayrıca plastitler bulunur. Bunların da kendi DNA'ları vardır ve endosimbiontlardan (bu durumda siyanobakteriler) oluşmuşlardır. Bu plastitlerin çoğu kloroplast olup siyanobakteriler gibi klorofil içerip fotosentez yoluyla enerji üretirler. Diğer plastitler gıda depolamaya yarar. Plastitlerin tek bir kökeni olması muhtemel olmakla beraber plastit bulunduran bitki grupları evrimsel olarak birbirine yakın değildir. Bazı ökaryotlar plastitlerini ikincil bir endosimbiyoz veya yutma yoluyla edinmişlerdir.
Çekirdeğin de endosimbiotik kökenli olduğu öne sürülmüş, ayrıntısı aşağıda verilmiştir. Ayrıca ökaryot kamçısının da endosimbiotik kökenli olduğu ve spiroketlerden geliştiği iddia edilmiş, ancak hücre anatomisi ile uyumsuzlukluğu ve hücre çoğalmasıyla bağdaştırılamadığı için bu iddia genel kabul görmemiştir.
Çoğu ökaryotun kamçı (flagella veya flagellum) adı verilen, ince, uzun ve hareketli sitoplazma uzantıları vardır. Bunlar tübülin ve kısa kirpiklerden (silyalardan) oluşur. Her ikisi de beslenme, duyum ve hareketle ilişkilidir ve prokaryot kamçılarından tamamen farklıdır. Kamçıyı içinden destekleyen bir demet mikrotüpçüktür. Bunlar, kinetozom ve sentriyol olarak da adlandırılan bazal cisimden çıkar. Mikrotüpçükler kamçının içinde, merkezde iki tekli ve onun çevresinde dokuz ikili olarak düzenlenmişlerdir. Kamçının ayrıca üzerini kaplayan saçlar (mastigonemler) ve onu zar ve hücre iskeletine bağlayan pullar da bulunabilir. Kamçının içi sitoplazmaya bağlıdır. Mikroiplik yapılar (aktin ve ona bağlanan α-aktinin, filamin, fimbrin) hücre zarının alt yüzeyinde tabaka ve desteler hâlinde bir ağ oluşturur. Mikrotübüllerin motor proteinleri, örneğin dinein ve kinesin, bu ağa dinamik bir özellik sağlar.
Sentriyoller çoğu zaman kamçısı olmayan hücre ve gruplarda da bulunur. Genelde kinetit olarak adlandırılan birli veya ikili gruplar hâlinde mikrotüpçüklerin köklerini oluştururlar. Bunlar hücre iskeletinin ana bileşkelerinden biridirler ve inşâları birkaç hücre bölünmesi boyunca olur. Bir kamçı ana hücrede kalır, öbürü ise ondan türetilir. Sentriyoller, ayrıca çekirdeğin bölünmesi sırasında iğ ipliği oluşumunda da rol oynayabilirler
Hücre iskeleti, hücrenin şeklini belirlemekte ve (kemokinez ve kemotaksi gibi) hareketini sağlamakta önemli bir rol oynar. Bazı protistlerin çeşitli başka mikrotüpçüklü organelleri de vardır: örneğin heliozoa ve radioloria, suda yükselme veya av yakalama için aksopodialara sahiptirler. Haptofitlerinse haptonema adlı kamçı benzeri bir organeli vardır.
Bitki hücrelerinin bir hücre duvarı vardır; bu, hücre zarının dışında yer alan ve pek esnemeyen bir tabakadır. Hem hücreye yapısal destek sağlar, hem de bir filtre mekanizması olarak çalışır. Hücrenin içine su girmesi hâlinde aşırı şişmeyi engeller. Birincil hücre duvarını oluşturan başlıca karbonhidratlar selüloz, hemiselüloz ve pektindir. Selüloz mikrofibrilleri, hemiselüloz bir yulara bağlanarak bir selüloz-hemiselüloz ağı oluştururlar. Bu da pektin matriks içinde yer alır. Birincil hücre duvarında bulunan başlıca hemiselüloz ksiloglukandır.
Çekirdek bölünmesi ile hücre bölünmesi çoğunlukla eşgüdümlü olur ve genelde mitoz yoluyla gerçekleşir. Bu süreç sonunda her kromozomun birer kopyası yavru hücrelere dağıtılır. Çoğu ökaryotlarda ayrıca bir eşeyli üreme süreci vardır. Bunda tipik olarak haploit ve diploit döller (nesiller) sırayla birbirini izler. Haploit dölde her kromozomun bir, diploit dölde ise iki kopyası vardır; haploit döl mayoz bölünmeyle, diploit dölse çekirdek kaynaşmasıyla (singami) meydana gelir. Ancak bu genel kalıp içinde epey bir çeşitlilik görülebilir.
Ökaryotik hücrenin oluşumu canlıların evriminde önemli bir dönem noktasıdır. Çünkü ökaryotlar tüm karmaşık hücreleri ve nerdeyse tüm çok hücreli canlıları içermektedir. Bu süreci oluşturan olayların zamanlamasını kestirmek zordur; Knoll (1992) yaklaşık 1,6-2,1 milyar yıl önce oluştuklarını öne sürer. Modern canlı gruplarına ait oldukları bâriz olan fosiller (kırmızı yosunlar), yaklaşık 1,2 milyar yıl önceden kalmadır.
1980'ler ve 1990'larda yapılan araştırmalar sonucunda çizilen rRNA Filogenetik ağaçlarında çoğu ökaryot organizma çözümlenememiş, (teknik anlamıyla gerçek taç olmayan) bir "taç" grup içinde bırakılmıştı. Bu grup, mitokondri kristallerinin şekline bağlı olarak ikiye bölünmüştü. Mitokondrisi olmayan birkaç grup, ayrı bir dal olarak diğer ökaryotlardan ayrılıyor, dolayısıyla bu eksikliğin çok eski dönemlerden kalma olduğu düşünülüyordu. Ancak artık bunun "uzun dal çekimi" adı verilen bir artifakt olduğu belirlenmiştir. Bu grupların diğer ökaryotlardan ayrıldıktan sonra mitokondrilerini kaybettikleri bilinmektedir.
Aktin ve diğer moleküllere dayandırılarak hazırlanmış ağaçlar, farklı ve daha tam bir görüntü ortaya çıkarmıştır. Ökaryotların çoğu aşağıdaki üst gruplara aittir:
Opisthokonta (Opistokontlar) Animalia (Hayvanlar), Fungi (Mantarlar), Choanoflagellatea (koanoflagellatlar), v. b. Amoebozoa (Amebozlar/Amebozolar) Çoğu kalın kökbacaklı amoeba (Amip) ve Mycetozoa (Sıvaşık küf) Rhizaria (Rizarlar) Foraminifera (Foraminiferler), Radiolaria (Radyolarlar) ve diğer çeşitli amipsi protozlar/protozolar Excavata (Ekskavatlar) Çeşitli kamçılı protozlar/protozolar Primoplantae/Archaeplastida (Primoplantlar/Arkeplastitler) Embryophyta (Kara bitkileri), yeşil algler, kırmızı algler ve Glaucophyta (Glaukofitler) Chromista (Kromistler) Kahverengi algler, diatomlar, Oomycete (su küfleri), v. s. Alveolata (Alveolatlar) Silyalılar, Apicomplexa (Apikompleksler), Dinoflagellata (Dinoflagellatlar), v. s.Birkaç sınıflandırma uzmanı, unikont ve bikontlar adlı iki büyük klad tanımaktadırlar: unikontlar tek, bikontlarsa iki kamçılı bir atasal canlıdan türemişlerdir. Bu sistemde ofistokontlar ve amibozoanlar unikont sayılırlar, kalanlar bikontturlar. Kromistler ve alveolatlar, atasal olarak kromalveolatlar adlı fotosentetik daha büyük bir gruba ait olabilirlerse de bu görüş hâlen tartışmalıdır. Özellikle sentrohelidler gibi bazı küçük protista grupları bu üst gruplardan hiçbiri ile ilişkilendirilmemişlerdir. Ökaryotlar, arkelerle genetik mekanizmaları açısından yakın derecede ilişkilidir. Bu yüzden bazı uzmanlarca arkelerle birlikte neomura kladına yerleştirilirler. Ancak hücre zarının yapısı gibi başka bakımlardan ökaryotlar bakterilerle benzeşirler. Bu durumu açıklamak için üç hipotez önerilmiştir:
Sonuncu hipotez halen en çok kabul görmektedir. İç zar sistemi ve mitokondrilerin kökeni de tartışmalıdır. Fagotrofik hipoteze göre endositozun gelişmesini takiben bu zarlar oluşmuş ve sonra özelleşmişlerdir. Mitokondriler, plastitler gibi yutma yoluyla edinilmişlerdir. Sintrofik hipoteze göre bir ön ökaryot (proto-ökaryot), beslenmek için ön mitokondriyi (proto-mitokondriyi) kullanıyordu. Zaman içinde büyüyerek onu tamamen sarmış, kısmen mitokondri genlerinin katkısıyla iç zarlar daha sonra oluşmuşlardır. (Bu hipotezin bir versiyonu hidrojen hipotezi olarak bilinir.)
Ökaryotlar (Latince: Eukaryota), hücrelerinde bir çekirdek ve başka organeller içeren bir canlılar grubu olup bilimsel sınıflandırmada arkeler ve bakterilerle beraber tüm canlıları kapsayan üç ana gruptan biridir.
Ökaryotların genetik malzemeleri zarla çevrili çoğunlukla bir, nadiren birden çok çekirdek içinde yer alırlar. Bu nedenle kelime, gerçek (Grekçe: eu) ve çekirdek (Grekçe: karyon) sözcüklerinden türetilmiştir. Sıfat hâli ökaryotiktir. Bakteri ve arkeler çekirdeksiz olduklarından beraberce prokaryot olarak adlandırılırlar (evvel (Grekçe: pro-) ve çekirdek (Grekçe: karyon)). Çekirdeğin yanı sıra ökaryotların kloroplast veya mitokondri gibi zarla çevrili çeşitli organelleri vardır. Bu tür hücre içi karmaşık yapılar da prokaryotlarda bulunmaz.
Ökaryotların ortak bir atası olduğu için bir üst âlem (İngilizce: domain) olarak tanımlanmışlardır. Üst âlem sisteminde ökaryotların prokaryotlara kıyasla arkelerle daha çok ortak özellikleri olduğundan arkelerle beraber neomura kladı içinde gruplandırılırlar.
Клітини прокаріотів (бактерій та архей) та еукаріотів на фенотипічному рівні схожі за наявністю ДНК та білок-синтезуючого апарату, представленого рибосомами; наявністю клітинних мембран; ферментних комплексів, що забезпечують процеси реплікації, транскрипції, трансляції та синтезу АТФ. До складу клітин прокаріотів та еукаріотів входять білки, жири, вуглеводи, нуклеїнові кислоти, мінеральні речовини та вода.
Відміни у будові клітин прокаріотів та еукаріотів пов'язані, в першу чергу, з тим, що майже вся ДНК прокаріотів складається з однієї хромосоми, і у випадку бактерій не пов'язана з білками-гістонами (хоча навіть бактерії мають гістоноподібні білки). Еукаріоти, на відміну від прокаріотів, зазвичай не містять генів, організованих у великі оперони, процес цитокінезу (поділу ДНК) проходить за участю веретена поділу. Рибосоми у еукаріотів більші, ніж рибосоми прокаріотів (умовна вага прокаріотичних рибосом становить 70S, тоді як еукаріотичних — 80S). Багато клітин еукаріотів здатні до фаго- та піноцитозу, мають морфологічно оформлене ядро, часто також мітохондрії, пластиди, ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, пероксисоми, мають іншу структуру джгутиків і джгутикових моторів, мають клітинний центр з центріолями. Для багатоьох клітин еукаріотів характерні мітоз, мейоз, повноцінний статевий процес, проте набагато нижча ймовірність горизонтального переносу генів. Зазвичай еукаріотичні клітини більші за розмірами, ніж прокаріотичні — їхній середній діаметр становить близько 5—20 мікрон, тоді як у бактерії та архей — 0,5—2 мікрона.
За палеонтологічними даними, прокаріоти на нашій планеті виникли близько 3,2 млрд років тому, тоді як еукаріоти є набагато молодшими — їх вік становить лише близько 1,6 млрд років[2]. Сьогодні в межах доменів бактерій та архей описано близько 40 тис. видів (хоча оцінки їх загальної кількості складають близько 10 млн видів[3]), тоді як у домені еукаріотів — близько 1,7 млн видів[2].
Першим відомим евкаріотом є наземний організм Diskagma buttonii, гіпотетичний вид грибів, рештки якого були знайдені у Південній Африці.[4]
Згідно з цією гіпотезою, еукаріотична клітина утворилась з прокаріотичної внаслідок розвитку плазмалемою системи інвагінацій, які надалі замкнулись навколо ділянок плазмалеми з ферментами дихального ланцюга, фотосинтетичними пігментами, нуклеоїда, відповідно утворивши мітохондрії, пластиди та ядро. Розвиток системи інвагінацій мембран призвів також до виникнення ендоплазматичної сітки, комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хлоропласти містять власну ДНК і розмножуються поділом, чому в мітохондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому поровий апарат мітохондрій та пластид подібний до порового апарату прокаріот, і відрізняється від порового апарату плазмалеми еукаріот, як виник мітоз та з'явились мікротрубочки, та ін.
Наприкінці XIX ст. видатний ботанік та фізіолог рослин М. С. Цвєт, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин можливо являють собою симбіотичний утвір з клітини-господаря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Це припущення у 20—40-х роках XX ст. було детально розроблено, в першу чергу, К. С. Мережковським у так звану ендосимбіотичну гіпотезу походження еукаріотичної клітини. Проте ідеї К. С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження еукаріотів називали вражаючим витвором людської фантазії", «луна-парком», «непристойною темою у колі освічених біологів»[2]. У 60-х роках гіпотеза ендосимбіотичного походження еукаріотичної клітини була заново сформульована американською дослідницею Л. Маргеліс.
Згідно з гіпотезою Мережковського-Маргеліс, еукаріотична клітина виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію, яка трансформувалась в мітохондрію. Далі клітина-господар, що містила мітохондрію, вступила в симбіоз з рухливою спірохетоподібною гетеротрофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, а потім його модифікація — мейоз, разом із статевим процесом. Виникла перша гетеротрофна еукаріотична клітина. Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. Симбіоз гетеротрофної еукаріотичної клітини з синьозеленою водорістю призвів до появи рослин, оскільки симбіотична синьозелена водорість надалі трансформувалась у пластиду.
Численні перевірки ендосимбіотичної гіпотези, виконані з широким використанням електронної мікроскопії, мікрохімічних та молекулярно-генетичних методів, з одного боку, підтвердили гіпотезу ендосимбіотичного походження мітохондрій та пластид, з іншого — автогенетичну гіпотезу походження ядра та більшості одномембранних клітинних органел. Таким чином, на початку XXI ст. теорія походження еукаріотів сформувалась на основі синтезу обох гіпотез — як автогенетичної, так і ендосимбіотичної, і отримала назву синтетичної гіпотези походження еукаріотів.
Сьогодні вважають, що першим кроком на шляху виникнення еукаріотів було утворення у гіпотетичного прокаріотичного пращура численних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкнули прокаріотичний нуклеоїд у двомембранну оболонку (тобто утворили морфологічно оформлене ядро), а з іншого — призвели до утворення ендоплазматичної сітки та похідного від неї комплексу Гольджі, а також травних вакуолей та їх похідних — лізосом.
На другому етапі, внаслідок горизонтального переносу гену, що кодує білок тубулін від спірохетоподібних бактерій, примітивні еукаріоти набули здатності до синтезу тубулінових мікротрубочок[2][5]. В результаті у еукаріотів з'явився цитоскелет, джгутики з базальним тілами, веретено поділу, мітоз. Базальні тіла джгутиків надалі у частини представників трансформувалися у клітинний центр, а порушення нормального мітозу (зокрема, скорочення інтерфази) призвели до виникнення мейозу і пов'язаного з ним статевого процесу. Група первинно безмітохондріальних еукаріотів сьогодні розглядається як перше, найпримітивніше царство еукаріотів — Hypochondria (Домітохондріальні, або Гіпохондріати)[6]. Всі гіпохондріати є одноклітинними гетеротрофами з тваринною стратегією живлення.
На третьому етапі еукаріотична клітина утворила симбіотичний комплекс з прокаріотичною клітиною, схожою із сучасними альфа-протеобактеріями. Ця прокаріотична клітина надалі трансформувалась у мітохондрію. За результатами як цитологічних, так і молекулярних досліджень, найдавнішими мітохондріальними еукаріотами вважаються ті, які мають мітохондрії з особливими дископодібними кристами. Група мітохондріальних еукаріотів з дископодібними кристами прийнята як таксон рангу царства — Discicristata (Дискокристати). В основі царства дискокристат знаходяться первинно гетеротрофні організми. На четвертому етапі еукаріоти розділилися на дві великі групи. Одна з цих груп має мітохондрії з трубчастими кристами, і складає царство Tubulocristates (Тубулокристати, трубчастокристні), друга — мітохондрії з переважно пластинчастими кристами, і приймається як царство Platycrsitates (платикристати, платівчастокристні). Подібно до дискокристат, в основі філ тубуло- та платикристат знаходяться первинно гетеротрофні організми із тваринною стратегією живлення.
На п'ятому етапі в еукаріотичному світі з'явилися перші рослини. За даними молекулярної та клітинної біології, ця подія пов'язана з симбіозом гетеротрофного еукаріота-платикристати з фотоавтотрофним прокаріотом — синьозеленою водорістю. Як наслідок цього симбіозу утворилась пластида, оточена двома мембранами, яка отримала назву первинно симбіотичної пластиди. Подальша дивергенція організмів з первинно симбіотичними пластидами обумовила виникнення у межах філи платикристат групи фотоавтотрофних відділів, які склали підцарство Plantae — рослини. Дивергенція гетеротрофних платикристат за типами живлення зумовила виникнення з одного боку, відділів з осмотрофним живленням — підцарства Fungi (гриби), з іншого — таксонів з фаготрофним живленням, що склали підцарство Animalia — тварини.
Велика радіація тубулокристат відбувалась майже паралельно з платикристатами, проте була пов'язана не стільки з еволюцією за типами живлення, скільки з еволюцією клітинних покривів та джгутикового апарату. В основі тубулокристат залишились амебоїдні організми, які складають підцарство Ameboflagellates (амебо-флогеляти). Дві прогресивніші філи представляють, з одного боку, таксони зі специфічними, альвеольованими покривами — Alveolata (альвеоляти), та з іншого — зі специфічними субмікроскопічними тричленними.
Первинні та вторинні ендосимбіози. Ендосимбіози, при яких в еукаріотичну клітину-хазяїна включався прокаріотичний симбіонт, що надалі трансформувався в клітинну органелу, називають первинними. На основі молекулярно-таксономічних досліджень наприкінці ХХ ст. було встановлено, що у світі еукаріотів первинні симбіози відбулися лише двічі. При першому ендосимбіозі прокаріотична бактерія, схожа з альфа-протеобактеріями, вступила в симбіоз з фаготрофним джгутиконосцем з царства гіпохондріат і трансформувалась у мітохондрію з трьома типами крист — дископодібними, трубчастими та пластинчастими. Подальша морфологічна еволюція мітохондрії відбувалася поступово, майже паралельно з клітиною-господарем. Так, у тубулокристат дископодібні кристи були редуковані, а збереглися трубчасті та інколи разом з ними — пластинчасті кристи. У платикристат були втрачені як дископодібні, так і трубчасті кристи, а збереглися лише пластинчасті (за винятком відділу Haptophyta).
Другий первинний ендосимбіоз відбувся між прокаріотичним оксигенним фотоавтотрофом — синьозеленою водорістю, та гетеротрофним організмом з царства платикристат. Як наслідок, виникла перша еукаріотична клітина, здатна до оксигенного фотосинтезу — так звана «протоводорість», тобто утворились рослини. У «протоводорості» пластида була вкрита двома мембранами, з яких внутрішня являла собою мембрану прокаріотичного ендосимбіонту — синьозеленої водорості, а зовнішня була похідною від плазмалеми клітини-господаря, і ймовірно являла собою видозмінену мембрану травної вакуолі, в яку потрапив ендосимбіонт.
«Протоводорість» дала початок трьом паралельним гілкам рослин з первинно симбіотичними пластидами — глаукоцистофітовим водоростям (Glaucocystophyta), червоним водоростям (Rhodophyta) та зеленим водоростям (Chlorophyta). Всі ці три відділи зберігають пластиди, що оточені лише двомембранною оболонкою. Глаукоцистофітові водорості зберігають цікаву атавістичну ознаку — між зовнішньою та внутрішньою мембраною пластиди розташовується шар муреїну — речовини, характерної для клітинних оболонок більшості евбактерій, і, зокрема, синьозелених водоростей. Відділ глаукоцистофітових є сліпою гілкою еволюції рослин.
Пластиди червоних водоростей — родопласти — також зберігають деякі яскраві ознаки спорідненості із синьозеленими водоростями, зокрема, особливі так звані фікобілінові пігменти. Пластиди зелених водоростей — хлоропласти — зберігають найменше фенотипічних рис схожості із прокаріотичним ендосимбіонтом. Від зелених водоростей беруть початок вищі рослини, причому всі вони зберігають первинно симбіотичні хлоропласти.
Червоні та зелені водорості неодноразово ставали ендосимбіонтами гетеротрофних еукаріотів з різних царств — дискокристат, тубулокристат та платикристат. Симбіози, що відбувалися за сценарієм «еукаріотичний хазяїн + еукаріотичний ендосимбіонт» називають вторинними. Внаслідок вторинних ендосимбіозів виникла велика група відділів водоростей із вторинно симбіотичними пластидами. Вторинно симбіотичні пластиди, на відміну від первинно симбіотичних, одягнені переважно чотирма мембранами, з яких внутрішня мембрана є залишком плазмалеми синьозеленої водорості (первинного ендосимбіонту), друга мембрана — видозміненою мембраною травної вакуолі господаря первинного ендосимбіонта (червоної або зеленої водорості), який є водночас вторинним ендосимбіонтом, третя мембрана — власне плазмалемою вторинного ендосимбіонта, четверта, зовнішня мембрана — плазмалемою господаря вторинного ендосимбіонта.
Цитоплазма, яка перебуває у проміжку між другою та третьою мембраною, так званий перипластидний простір, є редукованою цитоплазмою вторинного ендосимбіонта, і у деяких випадках зберігає 80S рибосоми та редуковану ендоплазматичну сітку. Зовнішні (третю та четверту) мембрани вторинно симбіотичних пластид називають хлоропластною ендоплазматичною сіткою.
Проте особливо цікавим є той факт, що у двох відділах водоростей із вторинно симбіотичними пластидами — хлорарахніофітових (Chlorarachniophyta) та криптофітових (Cryptophyta) — у перипластидному просторі зберігається навіть редуковане ядро вторинного ендосимбіонта — нуклеоморф. Нуклеоморф оточений двомембранною оболонкою і містить ДНК, яка по наявним у нуклеоморфі генам у хлорарахніофітових споріднена з ядерною ДНК зелених водоростей, а у криптофітових — з ядерною ДНК червоних водоростей.
У деяких водоростей, зокрема, у евгленофітових, вторинно симбіотичні пластиди вкриті не чотирма, а лише трьома мембранами. Вважається, що у цих водоростей одна з зовнішніх мембран пластиди була вторинно втрачена. Відомі випадки, коли пластиди (як первинно, так і вторинно симбіотичні) редукувалися, і тоді виникали вторинно гетеротрофні таксони. Зокрема, доведена вторинна редукція пластид у тварин з типу Apicomplexa, пращурами яких були фотоавтотрофні динофітові водорості (Dinophyta).
Є також дані, які дозволяють припустити, що оомікотові гриби (Oomycota) утворились внаслідок вторинного переходу до гетеротрофного типу живлення від однієї з груп водоростей-тубулокристат. Крім того, вторинно гетеротрофні таксони низьких та середніх рангів (від видів до порядків і навіть класів) відомі майже в кожному відділі водоростей, а також серед вищих рослин.
Поширення пластид через вторинні ендосимбіози обумовило виникнення значної кількості відділів водоростей. Наприклад, якщо водорості з первинно симбіотичними пластидами представлені лише трьома відділами (Glaucocystophyta, Rhodophyta, Chlorophyta), то водорості зі вторинно симбіотичними пластидами належать до 12 відділів. Серед них два відділи зберігають нуклеоморф (Chlorarachniophyta, Cryptophyta), 8 відділів мають чотиримембранні вторинно симбіотичні пластиди без нуклеоморфу, що походять від червоних водоростей (Raphidophyta, Chrysophyta, Eustigmatophyta, Xanthophyta, Phaeophyta, Bacillariophyta, Dictyochophyta, Haptophyta). Евгленофітові водорості мають тримембранні пластиди, пращурами яких є зелені водорості. У Dinophyta пластиди утворювались неодноразово, як внаслідок симбіозів з зеленими водоростями, так і навіть внаслідок третинних симбіозів зі вторинно симбіотичними фотоавтотрофами.
Поділ клітини відбувається безстатево мітозом, процес дозволяє кожному дочірньому ядру отримувати по одній копії кожної хромосоми. Цикл життя більшості еукаріотів включає сексуальну репродукцію, змінюючись між гаплоїдною фазою, де в кожній клітині присутня одна копія кожної хромосоми, та диплоїдною фазою, де в кожній клітині присутні по дві копії кожної хромосоми. Диплоїдна фаза являє собою злиття двох гаплоїдних гамет що формують зиготу, котра в свою чергу може ділитись в мітозі або пройти хромосомне скорочення в мейозі. В цьому шаблоні допустимі значні варіації — в тварин відсутня багатоклітинна гаплоїдна фаза, проте кожне покоління рослин може проходити через гаплоїдну та диплоїдну багатоклітинну фази.
В еукаріотів відношення площі до об'єму менше ніж в прокаріотів, тому метаболічний процес та процес утворення клітини триває довше.
Еволюція процесу сексуальної репродукції це первісна та фундаментальна характеристика еукаріотичних організмів. Базуючись на філогентичному аналізі, молекулярні біологи Дакс та Роджер припустили що статеве розмноження, хоч і не було основним способом розмноження, все ж було присвоїте спільному предку всіх еукаріотів. Основний комплект генів котрі фігурують на стадії мейозу присутній навіть в таких паразитичних видів як Trichomonas vaginalis та Giardia intestinalis, котрі до недавніх пір вважались асексуальними. Оскільки ці два види являються нащадками родів що на ранній стадії відділились від головного еволюційного дерева еукаріотів, було припущено що основні гени що фігурують в фазі мейозу, а отже і статеве розмноження, були швидше за все притаманними спільному предку еукаріотів. Еукаріотичні види що помилково вважалися безстатевими, наприклад паразит протозоя з роду Leishmania, все ж мають сексуальний цикл. Також, певні чинники вказують на те що амеба, котра вважалась безстатевим видом, з давніх часів розмножувалась статевим шляхом, та що в більшості сучасних безстатевих видів недавно та незалежно один від одного прокинувся сексуальний потяг.
Sinh vật nhân thực, còn gọi là sinh vật nhân chuẩn, sinh vật nhân điển hình hoặc sinh vật có nhân chính thức (danh pháp: Eukaryota hay Eukarya) là một sinh vật gồm các tế bào phức tạp, trong đó vật liệu di truyền được sắp đặt trong nhân có màng bao bọc. Eukaryote là chữ Latin có nghĩa là có nhân thật sự.
Sinh vật nhân thực gồm có động vật, thực vật và nấm - hầu hết chúng là sinh vật đa bào - cũng như các nhóm đa dạng khác được gọi chung là nguyên sinh vật (đa số là sinh vật đơn bào, bao gồm động vật nguyên sinh và thực vật nguyên sinh). Trái lại, các sinh vật khác, chẳng hạn như vi khuẩn, không có nhân và các cấu trúc tế bào phức tạp khác; những sinh vật như thế được gọi là sinh vật tiền nhân hoặc sinh vật nhân sơ (prokaryote). Sinh vật nhân thực có cùng một nguồn gốc và thường được xếp thành một siêu giới hoặc vực (domain).
Các sinh vật này thường lớn gấp 10 lần (về kích thước) so với sinh vật nhân sơ, do đó gấp khoảng 1000 lần về thể tích. Điểm khác biệt quan trọng giữa sinh vật nhân sơ và sinh vật nhân thực là tế bào nhân thực có các xoang tế bào được chia nhỏ do các lớp màng tế bào để thực hiện các hoạt động trao đổi chất riêng biệt. Trong đó, điều tiến bộ nhất là việc hình thành nhân tế bào có hệ thống màng riêng để bảo vệ các phân tử ADN (?) của tế bào. Tế bào sinh vật nhân thực thường có những cấu trúc chuyên biệt để tiến hành các chức năng nhất định, gọi là các bào quan. Các đặc trưng gồm:



Sinh vật nhân thực, còn gọi là sinh vật nhân chuẩn, sinh vật nhân điển hình hoặc sinh vật có nhân chính thức (danh pháp: Eukaryota hay Eukarya) là một sinh vật gồm các tế bào phức tạp, trong đó vật liệu di truyền được sắp đặt trong nhân có màng bao bọc. Eukaryote là chữ Latin có nghĩa là có nhân thật sự.
Sinh vật nhân thực gồm có động vật, thực vật và nấm - hầu hết chúng là sinh vật đa bào - cũng như các nhóm đa dạng khác được gọi chung là nguyên sinh vật (đa số là sinh vật đơn bào, bao gồm động vật nguyên sinh và thực vật nguyên sinh). Trái lại, các sinh vật khác, chẳng hạn như vi khuẩn, không có nhân và các cấu trúc tế bào phức tạp khác; những sinh vật như thế được gọi là sinh vật tiền nhân hoặc sinh vật nhân sơ (prokaryote). Sinh vật nhân thực có cùng một nguồn gốc và thường được xếp thành một siêu giới hoặc vực (domain).
Các sinh vật này thường lớn gấp 10 lần (về kích thước) so với sinh vật nhân sơ, do đó gấp khoảng 1000 lần về thể tích. Điểm khác biệt quan trọng giữa sinh vật nhân sơ và sinh vật nhân thực là tế bào nhân thực có các xoang tế bào được chia nhỏ do các lớp màng tế bào để thực hiện các hoạt động trao đổi chất riêng biệt. Trong đó, điều tiến bộ nhất là việc hình thành nhân tế bào có hệ thống màng riêng để bảo vệ các phân tử ADN (?) của tế bào. Tế bào sinh vật nhân thực thường có những cấu trúc chuyên biệt để tiến hành các chức năng nhất định, gọi là các bào quan. Các đặc trưng gồm:
Tế bào chất của sinh vật nhân thực thường không nhìn thấy những thể hạt như ở sinh vật nhân sơ vì rằng phần lớn ribosome của chúng được bám trên mạng lưới nội chất. Màng tế bào cũng có cấu trúc tương tự như ở sinh vật nhân sơ tuy nhiên thành phần cấu tạo chi tiết lại khác nhau một vài điểm nhỏ. Chỉ một số tế bào sinh vật nhân thực có thành tế bào. Vật chất di truyền trong tế bào sinh vật nhân thực thường gồm một hoặc một số phân tử ADN mạch thẳng, được cô đặc bởi các protein histone tạo nên cấu trúc nhiễm sắc thể. Mọi phân tử ADN được lưu giữ trong nhân tế bào với một lớp màng nhân bao bọc. Một số bào quan của sinh vật nhân thực có chứa ADN riêng. Một vài tế bào sinh vật nhân thực có thể di chuyển nhờ tiêm mao hoặc tiên mao. Những tiên mao thường có cấu trúc phức tạp hơn so với sinh vật nhân sơ.Эукарио́ты (устар. эвкарио́ты; лат. Eukaryota от др.-греч. εὖ- ‘хорошо’ или ’полностью’[2] + κάρυον ‘ядро’), или я́дерные, — домен (надцарство) живых организмов, клетки которых содержат ядро. Все организмы, кроме прокариот (бактерий и архей), являются ядерными. Вирусы и вироиды также не являются ни прокариотами, ни эукариотами; более того, сам вопрос, считать ли их живыми организмами, является дискуссионным.
Животные, растения, грибы, а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,6—2,1 млрд лет назад[3]. Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и поглощёнными этой клеткой бактериями — предшественниками митохондрий и пластид.
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органеллы (другое название, реже употребляемое в научной литературе[комм. 1], — органоиды), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств из-за биохимических особенностей. Также некоторые авторы выделяют царства протистов или простейших и хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалье-Смита, все эукариоты подразделяются на два монофилетических таксона (поддомена) — униконтов (Unikonta) и биконтов (Bikonta). Положение таких эукариот, как Collodictyon и Diphylleia rotans, на данный момент не определено.
Важнейшая, основополагающая особенность эукариот связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие, среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать достаточно прочную клеточную стенку из одной молекулы. Это обстоятельство могло заставить их использовать для клеточной стенки иной материал. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5—10 мкм, когда тот же показатель у эукариот составляет 10—100 мкм. Объём такой клетки в 1000—10 000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд лет назад, от них около 1,2 млрд лет назад произошли эукариотические организмы.
Комментарии:
Эукарио́ты (устар. эвкарио́ты; лат. Eukaryota от др.-греч. εὖ- ‘хорошо’ или ’полностью’ + κάρυον ‘ядро’), или я́дерные, — домен (надцарство) живых организмов, клетки которых содержат ядро. Все организмы, кроме прокариот (бактерий и архей), являются ядерными. Вирусы и вироиды также не являются ни прокариотами, ни эукариотами; более того, сам вопрос, считать ли их живыми организмами, является дискуссионным.
真核生物(学名:Eukaryota)是其细胞具有细胞核的单细胞生物和多细胞生物的总称,它包括所有动物、植物、真菌和其他具有由膜包裹着的复杂亚细胞结构的生物。
真核生物与原核生物的根本性区别是前者的细胞内含有细胞核,因此以真核来命名这一类细胞。许多真核细胞中还含有其它细胞器,如粒線體、叶绿体、高尔基体等。
由于具有细胞核,因此真核细胞的细胞分裂过程与没有细胞核的原核生物也大不相同。
真核生物在进化上是单源性的,都属于三域系统中的真核生物域,另外两个域为同属于原核生物的细菌和古菌。但由于真核生物与古菌在一些生化性质和基因相关性上具有一定相似性,因此有时也将这两者共同归于新壁總域演化支。
科學家相信,從基因證據來看,真核生物是細菌與古菌的基因融合體,它是某種古菌與細菌共生,異種結合的產物。
真核细胞之所以可以发展出有质膜包围的细胞核和细胞器,被认为有两个过程:膜内折和内共生。膜内折指始祖古核生物细胞的质膜内折成为真核细胞的膜系统。经过第二次内折,内质网膜形成高尔基体等细胞器。内共生学说指,古核生物吞入了需氧的、较小的、始祖原核生物(需氧真细菌,如变形细菌)和,由寄生过渡到共生,变为线粒体。通过类似的过程,古核生物细胞与较小的、始祖光合原核生物(如蓝细菌)共生,产生绿藻和红藻,第二次被古核生物吞入成为共生者,并演变为叶绿体。膜内折产生的原始的内质网膜系统,限制了吞入细菌的活动,避免了细胞成分被其“吃掉”。
在古时候,人们就已认识到动物和植物应分为两类。在林奈建立分类阶元时,虽然存在一定疑惑但还是把真菌划为两界说中的植物界。而后来的学者把真菌独立成一界,即把真核生物分为四界:原生生物界、植物界、动物界、真菌界。显微镜发明后,所发现的许多原生生物根据是自养还是异养,划分为原生植物与原生动物,分别归入植物界与动物界。而海克尔增加了原生生物界,即把真核生物分为四界。
在20世纪80年代之前,单细胞的原生生物的分类还处于很混乱的阶段,直到DNA测序技术的成熟使系统发生学出现,改变了这一状况,也使人们开始重新谱写真核生物的分类。
根据系统发生学的研究,根据进化分支上的同源性,2005年的国际原生生物学家协会(英语:International Society of Protistologists)提出了一种在当时被普遍接受的真核生物分类方法,分作六类(supergroups)。这种分类方法并没有明确这些类是生物分类学上的哪一等级,但其它文献将其当作"界".
古虫界 (Excavata) 鞭毛虫等原生生物 变形虫界 (Amoebozoa) 阿米巴与黏菌等 后鞭毛生物 (Opisthokonta) 动物、真菌、领鞭毛虫类 (choanoflagellate)等 有孔虫界 (Rhizaria) 有孔虫、放射虫、某些变形虫原生生物 囊泡藻界 (Chromalveolata) 不等鞭毛门 (Heterokonta)、定鞭藻门 (Haptophyta)、隐藻门 (Cryptophyta) , 囊泡虫总门 (Alveolata) 泛植物界 (Archaeplastida) 陆生植物、绿藻、红藻、灰胞藻但是,这一"六界说"很快就遭到了新的研究的挑战,特别是对囊泡藻界是否有共同起源的质疑。真核生物的分类目前仍有很大的不确定性。例如,把真核生物分为单鞭毛生物(Unikonta)与双鞭毛生物(Bikonta),前者包括变形虫界与后鞭毛生物,后者其它四界。但是,很多研究表明单鞭毛生物与双鞭毛生物都不是同源的进化支。另外,有孔虫界应该并入囊泡藻界。泛植物界也应该不是同源的,需要被细分。
一些研究拆分了囊泡藻界,并将其一部分与有孔虫界合并为SAR超类群,另一部分视为原始色素体生物的近缘类群 ,比如Burki等在2007提出的系统树
真核生物另一些研究则认为Hacrobia和古虫界的一部分应该属于原始色素体生物,和SAR超类群组成双鞭毛生物,变形虫和后鞭毛生物组成单鞭毛生物:
真核生物 双鞭毛生物原始色素体生物 + Hacrobia + 古虫界(除了Preaxostyla)
Preaxostyla (原本属于古虫界)
SAR = 不等鞭毛类 + 囊泡虫类 + 有孔虫界
单鞭毛生物变形虫界
后鞭毛生物
还有一些研究认为囊泡藻界不应被拆分,而是将SAR超类群归入扩大的囊泡藻界,如Rogozin 等在2009年的一项研究:
真核生物原始色素体生物
古虫界(位置未定)
单鞭毛生物变形虫界
后鞭毛生物
许多观点将扩大的囊泡藻界视为原始色素体生物的近缘类群:
真核生物古虫界
原始色素体生物
单鞭毛生物变形虫界
后鞭毛生物
2009年一项研究详细研究了已有的真核生物分类的证据,提出了如下分类:
真核生物2012年的一项研究也支持单鞭毛生物与双鞭毛生物的划分:
真核生物 双鞭毛生物Hacrobia
SAR = 不等鞭毛类 + 囊泡虫类 + 有孔虫界
单鞭毛生物现在广泛认为有孔虫和不等鞭毛类及囊泡虫类是姊妹类群,同属于一个超类群,因此有孔虫也不再视为真核生物的主要类群之一,同时单鞭毛生物及其子类群变形虫和后鞭毛生物都被视为单系群,即动物和真菌同属一个单系群--后鞭毛生物。除此之外,还没有其它共识。
真核细胞体积通常远大于原核细胞,其内部有各种内膜结构构成的内膜系统。细胞器与微丝、微管、中间丝等细胞骨架在细胞的功能和结构上起到了重要的作用。真核DNA线性地划分在几个染色体上,平时以染色质形态存在于细胞核中,在有丝分裂时由微管组成的纺锤丝分离。
真核细胞包含多种膜结构,统称为内膜系统。膜可以部分脱离,产出独立的小室,称为小泡或液泡。很多细胞通过胞吞作用摄入食物和其他物质,外层膜首先入鞘,然后收紧形成独立的小泡弹出。大多其他由膜包围的细胞器都可能形成自这种液泡。
细胞核由两层膜包围,常称为核膜,包含核孔,使得物质能够进出。核膜的管状、片状突出形成了内质网,负责蛋白质的生成和转运。粗面内质网上有核糖体,进行蛋白质合成,这些蛋白质随后进入细胞内腔,再进入小泡中,然后从光面内质网弹出。对于大多真核细胞,这些携带蛋白质的小泡释放到一堆扁平囊泡中,即高尔基体,并进一步改装。
小泡可以被特化,用于多种不同目的。例如,溶酶体装有可以分解食物的酶,過氧化物酶用于分解有毒的過氧化物。很多原生生物有可收缩的液泡,用于收集和排出多余的水,排出小体(英语:extrusomes)可以喷出物质,用于击偏捕食者,或抓住猎物。而在高等植物中,中央液泡占据细胞多半的体积,该液泡也维持了它的渗透压。
粒线体是几乎所有真核细胞共有的细胞器,有两层膜,即两层磷脂双分子层所包裹,内膜向内折叠成嵴,在此进行有氧呼吸。线粒体有自己的DNA,今天通常认为是由内共生学说的原核生物发展而来,很可能是接近立克次體的變形菌門細菌。少数原生生物中没有线粒体,却发现具有由线粒体起源的细胞器,如氢化酶体和纺锤剩体(英语:Mitosome),因此很可能这些原生生物是在之后才丢失了线粒体。
植物和很多藻类拥有细胞器色素体。同样的,这些色素体也有自己的DNA,起源于内共生,这次的内共生体是藍菌門细菌。色素体又称质体,最常见的就是叶绿体,像蓝菌门细菌一样也有叶绿素,通过光合作用合成有机物(如葡萄糖)。其他色素体参与储存食料。虽然色素体很可能是单起源的,但并非所有含有色素体的细胞也是单起源的,实际就有些真核细胞通过多重内共生或摄入获得色素体。
内共生起源也许能适用于细胞核,此前鞭毛也被认为起源于螺旋體。但由于缺乏细胞生物学证据,且难以与细胞繁殖过程调和,螺旋體一说未被广泛接受。
许多真核生物具有细长且能运动的突起,称为鞭毛,以及较短但类似的结构—纤毛,合称为波动足(英语:undulipodia),与真核生物的运动、感觉和摄食密切相关,主要成分是微管蛋白。这些细胞器与原生生物的鞭毛具有鲜明的不同,基体伸出一簇微管,支撑起它们。基体也被称作毛基体或中心粒,是中心体的主要组成部分。鞭毛本身也可以再有茸毛,以及鳞片,连接膜和内杆,它们又与细胞质相连。
微丝结构由肌动蛋白和结合了肌动蛋白的蛋白质构成,例如α-肌动蛋白、丝束蛋白(英语:fimbrin)和细丝蛋白(英语:filamin)也存在于膜下皮质层和皮质束众。微管中的马达蛋白,如动力蛋白,或驱动蛋白和肌动蛋白,如动力蛋白,为系统提供了动态特性。 即使是没有鞭毛的细胞也常有中心粒,但开花植物和松柏门植物则没有。中心粒通常成组出现,称作动胞器(kinetids),形成多种不同的微管根,它们构成了细胞骨架结构的主要成分,往往在多次细胞分裂的过程中合成,一个鞭毛从亲代保留下来,另一个则据亲代衍生而来。此外,中心粒也可能与细胞核分裂时纺锤体的形成有关。
以有丝分裂的形式,细胞核的分裂通常与细胞分裂得以相互协调,这一过程使得每一个子代细胞核都得到亲代的一条染色体拷贝。在大多真核细胞中,还有另一种有性繁殖过程,即减数分裂,这一过程中,二倍体亲代细胞经由两次分裂成为单倍体,DNA的数量减半。然而,减数分裂本身也分很多种类。[1]
相比原核细胞,真核细胞有更小的表面积-体积比,因此新陈代谢速率较慢,细胞周期更长。在有些多细胞生物体中,一些专门用于新陈代谢的细胞会扩大表面积,如小肠绒毛细胞。
现在的真核生物广泛地采用有性生殖,并有证据显示,这是真核生物原始、根本的特性。基于系统发生学分析,生物学家达克斯和罗杰提出,真核细胞共有的祖先进行随机性行为。减数分裂中核心的一组基因出现在阴道毛滴虫(英语:Trichomonas vaginalis)和肠贾第虫中,而这两种生物此前被视为是无性的。在真核细胞进化树上,这两个物种很早便分离出来,因此可以推断,减数分裂的核心基因存在于所有真核生物的那个共同祖先中,也因此这一祖先是有性的。其他对真核物种的研究也揭露了生殖周期的证据。例如寄生原生生物利什曼原蟲近来显示存在生殖周期。证据显示,此前被认为是无性的阿米巴虫,在古代也是有性的,并且如今大多无性生物也是近期才独立地进化为无性的。
植物细胞也与其它真核细胞有很多不同。其特征是:
真菌细胞与动物细胞最为相似,但存在以下不同:
动物细胞是组成动物组织的基本单元。动物细胞与其它真核细胞有巨大差异,主要体现在缺少细胞壁、大液泡等,但具有能进行胞吞、胞吐的小液泡。也正是因为缺乏细胞壁,动物细胞可以转换成不同的形态,吞噬細胞正是具有这点才能进行吞噬作用。
动物细胞可以分化成许多种类型,比如成人体内大约有210种细胞。
真核生物(学名:Eukaryota)是其细胞具有细胞核的单细胞生物和多细胞生物的总称,它包括所有动物、植物、真菌和其他具有由膜包裹着的复杂亚细胞结构的生物。
真核生物与原核生物的根本性区别是前者的细胞内含有细胞核,因此以真核来命名这一类细胞。许多真核细胞中还含有其它细胞器,如粒線體、叶绿体、高尔基体等。
由于具有细胞核,因此真核细胞的细胞分裂过程与没有细胞核的原核生物也大不相同。
真核生物在进化上是单源性的,都属于三域系统中的真核生物域,另外两个域为同属于原核生物的细菌和古菌。但由于真核生物与古菌在一些生化性质和基因相关性上具有一定相似性,因此有时也将这两者共同归于新壁總域演化支。
科學家相信,從基因證據來看,真核生物是細菌與古菌的基因融合體,它是某種古菌與細菌共生,異種結合的產物。
 分類 ドメ
分類 ドメEukarya
和名 真核生物 英名 Eukaryote クラスター[1]真核生物(しんかくせいぶつ、学名: Eukaryota、英: Eukaryote)は、動物、植物、菌類、原生生物など、身体を構成する細胞の中に細胞核と呼ばれる細胞小器官を有する生物である。真核生物以外の生物は原核生物と呼ばれる。
生物を基本的な遺伝の仕組みや生化学的性質を元に分類する3ドメイン説では、古細菌(アーキア)ドメイン、真正細菌(バクテリア)ドメインと共に生物界を3分する。他の2つのドメインに比べ、非常に大型で形態的に多様性に富むという特徴を持つ。かつての5界説では、動物界、植物界、菌界、原生生物界の4界が真核生物に含まれる。
真核生物の細胞は一般的に原核生物の細胞よりも大きく、場合によっては1000倍以上の体積を持つこともある。細胞内にはさまざまな細胞小器官がある。細胞核は必要な物質のみ透過する穴の開いた二重の膜で覆われており、核液と遺伝情報を保持する DNA を含んでいる。細胞のその他の部分は細胞質と呼ばれ、細胞骨格によって支えられている。
核の周囲を板状とチューブ状の小胞体が取り巻いている。チューブ状の物は滑面小胞体、板状の物は粗面小胞体と呼ばれており、粗面小胞体にはいくつものリボソームが張り付き、細胞内での物質の生成、伝達が原核生物と同程度に潤滑させる器官である。リボソーム内で合成されたタンパク質が小胞体に渡され、小胞に入れられて細胞全体に分配される。ほとんどの真核生物では小胞はゴルジ体に蓄積される。小胞には様々な種類があり、これらの動きを合わせて細胞内組織が構成されている。
その他にも多くの器官が存在している。原核細胞と異なり、真核細胞の中には異化作用と酸素の消費に関係するミトコンドリアがある。植物や藻の系列では細胞内に光合成を行う葉緑体も含まれている。葉緑体を内部に持つ原核生物も存在しているが、それぞれ別々に細胞内に取り込まれたと考えられている(細胞内共生説)。真核生物の多くは細胞表面に鞭毛や繊毛があり、移動に使用したり、あるいは受容器官の働きをしている物もある。
細胞分裂の際には、まず核分裂が行われる。一般に、核内のDNAは細胞分裂に先立って、より集められて染色体になり、DNAはこの染色体ごとに新しい2つの細胞のために糸状の構造(紡錘糸)によって分かれて運ばれる。分かれた染色体のコピーが渡される。これを有糸分裂と呼ぶ。真核生物の染色体は直鎖状であり、末端にはテロメアと呼ばれる構造がある。
ほとんどの真核生物では有性生殖が行われる。減数分裂後、染色体の半数体を2つ合わせて核の合成を行う。これには様々なパターンが存在する。
真核生物の本体は進化論的には古細菌の姉妹群、または古細菌そのもの(エオサイト説)とする説が有力である。これは転写や翻訳など遺伝情報に関わる遺伝子、一部の細胞質で働く遺伝子が古細菌に近いことから支持されている。一方でミトコンドリアや葉緑体で働く遺伝子や一部の代謝に関連する遺伝子は真正細菌と類似性を示している。これは遺伝子の水平伝播および共生したミトコンドリアや葉緑体などに由来する。成立過程は捕食説(αプロテオバクテリアと大型の古細菌がもう一方を捕食しようと試みたとする説)、水素説(メタン生成古細菌とαプロテオバクテリアによる廃棄物のやり取りをもとにした共生)などいくつか説があるが、いずれにせよ20億年前までには真核生物が成立したと考えられている。
最も古い真核生物の化石は、27億年前の地層から検出されたステランと呼ばれる真核生物に由来する有機物質である。また、真核生物の化石そのものも21億年前の地層から発見されている。ただし、これらの化石が真に真核生物由来かどうか、なお議論の必要がある。
分子系統以前に広く受け入れられていた五界説では、真核生物に含まれるのは、動物、植物、菌類、原生生物といった4つの界(と呼ばれるまとまり)である。しかし、近年では、ゲノム解析などの研究成果を受けて新しい分類法が次々と発表されている。例えば、動物と菌類は同一の系統に含まれるとして、オピストコンタとして上位分類を設けている。
以下に国際原生生物学会による公式的な分類体系(Adl et al. 2012)の概観を表に示した[1]。
ドメイン cluster super-group first rank 例 真正細菌 古細菌 真核生物 アモルフェアこれは全真核生物を、オピストコンタ、アメーボゾア、エクスカバータ、アーケプラスチダ、SAR の5つのスーパーグループにまとめる。ただし、正確には、スーパーグループが不明な小さな系統が多数ある。
スーパーグループのうち、オピストコンタとアメーボゾア、アーケプラスチダとSARは、それぞれ、クラスターにまとめられる(すなわち真核生物は大きく3つに分かれる)。スーパーグループ不明の系統のいくつかも、系統的に内包される場合はクラスターに含まれるが、正確な系統が不明なためにどれが含まれるかの詳細は不明である。
ISOPの分類の前の版である2005年の分類では、SARの代わりにクロモアルベオラータ(ストラメノパイル + アルベオラータ + クリプト藻 + ハプト藻)とリザリアがスーパーグループとなっており、計6スーパーグルプがあった。クラスターの概念はなかった。
表に示したように、動物や植物は、原生生物の分類体系内に収まっている。これ以前の原生動物の分類体系には、国際原生生物学会の前身である国際原生動物学会による合意体系としてLevine et al. (1980) があり、図鑑や百科事典で見られる分類体系の多くはこれに従っている。
真核生物(しんかくせいぶつ、学名: Eukaryota、英: Eukaryote)は、動物、植物、菌類、原生生物など、身体を構成する細胞の中に細胞核と呼ばれる細胞小器官を有する生物である。真核生物以外の生物は原核生物と呼ばれる。
生物を基本的な遺伝の仕組みや生化学的性質を元に分類する3ドメイン説では、古細菌(アーキア)ドメイン、真正細菌(バクテリア)ドメインと共に生物界を3分する。他の2つのドメインに比べ、非常に大型で形態的に多様性に富むという特徴を持つ。かつての5界説では、動物界、植物界、菌界、原生生物界の4界が真核生物に含まれる。
진핵생물(眞核生物, 영어: Eukaryota)은 진핵세포를 가진 생물을 말한다. 진핵세포(眞核細胞)는 세포 내에 핵으로 대표되는 다양한 세포소기관을 가진 세포들을 통칭하는 이름이다. 세포소기관 중 미토콘드리아나 엽록체는 과거 진화 과정에서 외부의 원핵세포들이 세포내 공생체로 도입된 경우로 생각되며, 그 외의 소기관들도 각각 특정한 업무를 수행하도록 분화되어 있다. 원핵세포의 DNA와는 달리, 진핵세포의 DNA에는 필요 없는 부분(인트론)이 대단히 많이 존재하며, 따라서 필요한 부분(엑손)을 이어서 mRNA를 만들게 된다. 또한 리보솜은 원핵세포보다 큰 80S이다.
진핵생물은 단계통군으로 나타나며, 3개의 생물 도메 중의 하나를 구성한다. 나머지 2개의 도메인은 세균과 고균으로, 원핵생물로 불리며 이상과 같은 특징을 갖고 있지 않다. 진핵생물은 모든 생명체 중에서는 일종의 소수이다. 심지어 인체에는 사람의 세포수보다 10배나 많은 미생물이 들어 있다.[3]그러나 진핵생물은 계 수가 가장많다. 3개의 역 중에 거의 가장 많은 종이 알려져 있다.
다음은 2006년 토머스 캐빌리어스미스가 제안한 생명의 나무 계통 분류이다.[4]
LUA 스핑고박테리아 네오무라다음은 아들(Adl et al. 2012)과 부르키(Burki, 2014/2016) 등의 진핵생물 계통 분류이다.[5][6][7]
진핵생물 AH/SAR 원시색소체생물진핵생물(眞核生物, 영어: Eukaryota)은 진핵세포를 가진 생물을 말한다. 진핵세포(眞核細胞)는 세포 내에 핵으로 대표되는 다양한 세포소기관을 가진 세포들을 통칭하는 이름이다. 세포소기관 중 미토콘드리아나 엽록체는 과거 진화 과정에서 외부의 원핵세포들이 세포내 공생체로 도입된 경우로 생각되며, 그 외의 소기관들도 각각 특정한 업무를 수행하도록 분화되어 있다. 원핵세포의 DNA와는 달리, 진핵세포의 DNA에는 필요 없는 부분(인트론)이 대단히 많이 존재하며, 따라서 필요한 부분(엑손)을 이어서 mRNA를 만들게 된다. 또한 리보솜은 원핵세포보다 큰 80S이다.
진핵생물은 단계통군으로 나타나며, 3개의 생물 도메 중의 하나를 구성한다. 나머지 2개의 도메인은 세균과 고균으로, 원핵생물로 불리며 이상과 같은 특징을 갖고 있지 않다. 진핵생물은 모든 생명체 중에서는 일종의 소수이다. 심지어 인체에는 사람의 세포수보다 10배나 많은 미생물이 들어 있다.그러나 진핵생물은 계 수가 가장많다. 3개의 역 중에 거의 가장 많은 종이 알려져 있다.


















































.svg)





.jpeg)
.jpg)















































































{kind=link}
{kind=link}
{kind=link}
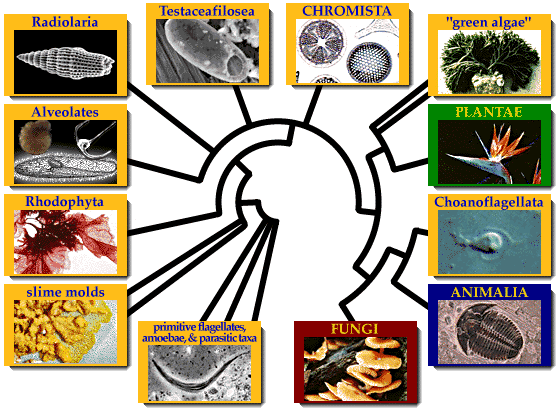{kind=link}