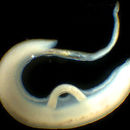
Els digeneus (Digenea, gr. dis, "doble" i genos, "raça") són una subclasse de platelmints de la classe dels trematodes. Es tracta de cucs paràsits amb un tegument sincitial, en general proveïts de dues ventoses, una oral i una altra ventral. Els adults parasiten especialment el tub digestiu, però poden infestar qualsevol òrgan de totes les classes de vertebrats, inclòs l'ésser humà. Fasciola hepatica produeix la fasciolosis, un malaltia parasitària dels remugants que pot afectar a l'home. Antigament els hi relacionava amb els Monogenea, però en l'actualitat s'admet que són més propers als Aspidogastrea i que els Monogenea estan més relacionats amb els Cestoda. S'han descrit unes 6.000 espècies.
Digenea (grč. dis – dvostruko, genos – pleme) je razred reda Trematoda u koljenu Platyhelminthes . Sastoji se od parazitskih pljosnatih glista s (poznatog kao metilji ) sa sincicijskim tegumentom (tjelesnim omotačem) i obično, dvije pijavke, jedna ventralna i jedna usna. Odrasli obično žive u probavnom traktu, ali se javljaju u svim organima svih razreda kičmenjaka. Nekada se mislilo da su povezane sa Monogenea, ali je prepoznato da su one najbliže grupi Aspidogastrea i da su monogenee bliže Cestoda. Do danas je opisano oko 6 000 vrsta.
Karakteristična obilježja Digenea uključuju sincicijski tegument; to jest tegument gdje su spojevi između sćelija razgrađeni pa jedna kontinuirana citoplazma okružuje cijelu životinju. Sličan tegument nalazi se i u ostalim članovima Neodermata; grupa Platyhelminthes uključuje Digenea, Aspidogastrea, Monogenea i Cestoda. Digenea imajuju crvoliko, nesegmentirano tijelo i čvrst parenhim, bez tjelesne šupljunešupljine (celom)a kao kod svih pljosnatih glista.
Obično postoje dva pijavke, prednja oralna I acetabularna, na ventralnoj površini. Oralna okružuje usta, dok je ventralna pijavka slijepi mišićni organ koji nema veze s bilo kojom unutrašnjom strukturom. Monostome su crvi s jednom pijavkom (oralnom). Trematoda sa oralnom i acetabulumom pijavkom na zadnjem kraju tijela nazivaju se Amfistomi". Distome su oblici sa oralnim i ventralnim sisaljkama, ali ventralni je negdje drugdje osim otraga. Ovi su pojmovi uobičajeni u starijoj literaturi, kada se smatralo da odražavaju sistemske odnose unutar grupa. U savremenoj taksonomiji Digenea, nisu više u upotrebi.
Ogromna većina Digenea su hermafroditi . To je vjerovatno prilagođavanje maloj brojnosti unutar domaćina, omogućavajući da se životni ciklus nastavi i kada samo jedna individual uspješno zarazi krajnjeg domaćina. Oplodnja je unutrašnja, a sperma se prenosi cirusom u Laurerov kanal ili genitalni otvor. Ključna grupa Digenea koje su dvodomne su šistosome. Sveprisutna je seksualna reprodukcija u prvom larvalnom stadiju . Dok se seksualna formacija jaja Digenea i aseksualna reprodukcija u prvom stadiju larve (miracidij) široko izvještavaju, razvojna biologija aseksualnih faza ostaje problem. Studije elektronskim mikroskopskom pokazale su da se svetlosnomikroskopski vidljive kuglice zametaka sastoje od ćelija u mitozi, koje stvaraju embrion i liniju novih zametnih ćelija kojie ostaju uključene u ove embrionske faze. Budući da izostanak mejotskih procesa nije dokazan, tačna definicija ostaje dvojbena.
Protandrija je opće pravilo kod Digenea. Obično su prisutna dva testisa, ali neke mogu imati i više od 100. Prisutni su i vasa eferentia, vas deferens, sjemenska vezikula, ejakulatorni kanal i cirus (analogan penisu) koji je obično (ali ne uvijek) zatvoren u vrećicu. Cirus se može, ali ne mora prekrivati i bjelančevinskim bodljicama. Tačna konformacija ovih organa unutar genitalija muških terminala je taksonomski važna na porodičnoj i generičkoj razini.
Obično postoji jedan jajnik sa jajovodom , semenska posuda, par vitelinskih žlijezda (uključenih u stvaranje žumanjka i proizvodnju ljuske jajeta) s kanalima, otitip (komora u kojoj se formiraju jajašca), složena kolekcija žljezdanih ćelija koja se naziva Mehlisova žlijezda' ', za koju se veruje da podmazuju matericu za prolazak jajašca.
Osim toga, neke digenea pimaju kanal zvan Laurelovog, koji vodi od ovidukta do dorzalne površine tijela. Raspravlja se o funkciji ovog kanala, ali može se koristiti za osjemenjavanje kod nekih vrsta ili za odlaganje otpadnih proizvoda reprodukcije u drugim vrstama. Većina trematoda posjeduje ovikapt, povećani dio jajovoda koji se pridružuje jajniku. Vjerovatno kontrolira oslobađanje iz jajnika i raširuje se prema dolje u maternicu. Maternica se obično otvara u zajednički genitalni atrij koji je prima distalni muški kopulatorni organ (cirus) prije nego što se otvori na vanjskoj površini crva. Distalni dio maternice može se proširiti u metratermu, koja se mišićnim sfinkterom odvaja od proksimalnog diela maternice ili se može postaviti uz bodlje, kao u Monorchiidae i nekim drugim porodicama.
Kao odrasli, većina Digenea ima terminalna ili subterminalna usta, mišićno ždrijelo koji daje snagu za gutanje hrane, i razranat, slijepi probavni sistem koji se sastoji od dvije cjevaste vreće zvane ceka (sing. cekum). Kod nekih vrsta dvije crijevne ceke pridružuju se posredno i čine crijevo u obliku prstena ili ciklocel. U ostalih se ceke mogu stopiti sa stražnjim tjelesnom zidom da bi se stvorio jedan ili više anusa ili sa ekskretornom vezikulom da bi se formirao uroprokt. Digene su također u mogućnosti za direktno usvajaju hranjive tvari kroz tegument pomoću pinocitoza i fagocitoza u sinciciju. Većina odraslih javlja se u probavn om traktu kičmenjaka ili pridruženih organa), gdje se najčešće uzima sadržaj lumena (npr. hrana koju je unio domaćin, žuč, [[sluz) , ali mogu se hraniti i kroz zid sluznice (npr. submukoza, krv domaćina). Krvni fluktuiraju, poput šistosoma, spirorhiida i sanguinicolida, a hrane se isključivo krvlju. Aseksualne faze u mehkušcima, srednjim domaćinima, hrane se većinom direktnom apsorpcijom, iako stadij larve redija koji se nalazi u nekim skupinama ima usta, ždrijelo i jednostavno crijevo, pa može aktivno trošiti tkivo domaćina ili čak i drugo parazitirati. Incestirani metacerkarijski stadiji i slobodno živući cerkarijski lstadiji se ne hrane.
Uparene ganglije na prednjem kraju tijela služe kao mozak. Od toga se živci protežu sprijeda i straga. Senzorni receptori uglavnom nedostaju među odraslima, iako imaju tangoreceptorske ćelije. Stadiji larve imaju mnogo vrsta senzornih receptora, uključujući one za svjetlost i hemoreceptore. Hemorecepcija ima važnu ulogu u miracidijskoj larvi koja slobodno živi i prepoznaje i locira svog domaćina.
Postoji složen niz varijacija u složenom životnom ciklusu Digenea, a plastičnost u ovoj sposobnosti vjerovatno je ključna za uspjeh grupe. Općenito, životni ciklusi mogu imati dva, tri ili četiri obvezna (potrebna) domaćina, ponekad s prijenosom ili paratenijskim međudomaćinima . Životni ciklus s tri domaćina vjerovatno je najčešći. U gotovo svim vrstama prvi domaćin u životnom ciklusu su mehkušci.[2] Ovo je dovelo do zaključka da je predak bio parazit mehkušaca i da su domaćini kičmenjaci dodani naknadno. Izmjena seksualnih i aseksualnih generacija važno je obilježje Digenea. Ovaj fenomen uključuje prisustvo nekoliko diskretnih generacija u jednom životnom ciklusu.
Tipski životni ciklus digenanskih trematoda je sljedeći. Jaja napuštaju kičmenjačkog domaćina u izmetu i koriste različite strategije da zaraze prvog srednjeg domaćina, u kojem se ne događa seksualna reprodukcija. Digenea mogu zaraziti prvog srednjeg domaćina (obično puža) bilo pasivnim ili aktivnim sredstvima. Naprimjer, jaja nekih od njih (pasivno) jedu puževi (ili, rijetko, crvi annelida) u kojima se izlegu. Alternativno, jaja se mogu odlagati u vodu da bi oslobodila larvu koja pliva, trapljasti miracidij, koji mora locirati i prodrijeti u tjelesni zid domaćina puža.
Nakon incestacije puževa ili prodora u puža, miracidij metamorfozira u jednostavnu, majčinsku sporocistu sličnu vrećici. Matična sporocista prolazi kroz krug unutarnjih b espolnih dijeljenja, što stvara ili „redije“ ili „kćeri sporociste“. Druga generacija je, dakle, kći parhenitske sekvence. Oni prolaze daljnju aseksualnu reprodukciju, što u konačnici rezultira velikim brojem drugog stadija slobodno živuće larve zvane cercarija.
Cerkarije koje plivaju slobodno napuštaju domaćiske puževe i kreću se kroz slatkovodnu ili morsku okolinu, često koristeći rep poput biča, iako postoji ogromna raznolikost morfologije repa. Cercarije su zarazne drugom domaćinu u životnom ciklusu, a infekcija se može dogoditi pasivno (npr. ribe) konzumiranjem cerkarija ili aktivno (cerkarija prodire kroz ribu).
Životni ciklusi nekih vrsta uključuju samo dva domaćina, a drugi je kičmenjak. U tim skupinama seksualna zrelost se događa nakon što cerkarija probije tjelesni omotač drugiog domaćin, što je u ovom slučaju i definitivni domaćin. Životni ciklusi sa dva domaćina mogu biti primarni (nikad nije postojao treći domaćin) kao u Bivesiculidae ili sekundarni (postojao je svojedobno u evolucijskoj historiji treći domaćin, ali je izgubljen).
U životnim ciklusima s tri domaćina, cerckarije se u drugom intermedijarnom domaćinu razvijaju u stadiju mirovanja, "metacerkarija" koja se obično učahuri u cistu domaćina ili je kapsulirana u sloj tkiva izvedenog samo od domaćina. Ova faza je infektivna za definitivnog domaćina. Prijenos se događa kada je konačni domaćin predator inficiranog drugog srednjeg domaćina. Metacerkarije se ekscistiraju u crijevima konačnog domaćina kao odgovor na različite fizičke i hemijske signale, kao što su nivoi pHu [u krvi], probavni enzimi], temperatura itd. Jednom kada su ekscistirani, odrasli migriraju na manje ili više specifična mjesta u konačnom domaćinu i životni se ciklus ponavlja.
O evolucijskom porijeku Digenee raspravlja se već neko vrijeme, ali čini se da se općenito slaže da je protodigenea bio parazit mehkušaca, moguće u plaštanoj šupljini. Dokaz za to proizlazi iz sveprisutnosti mehkušaca kao njihovih prvih posrednih domaćina, te činjenice da većina aspidogastreanskih oblika (sestrinska grupa Digenei) također ima zjednice sa mehkušacima. Smatra se da su rani trematodi (skupni naziv za Digenea i Aspidogastrea) vjerovatno evoluirali iz rabodokolskih turbellaria koji su kolonizirali otvorenu šupljinu plašta ranih mehkušaca. Vjerojatno su se složeniji životni ciklusi razvijali putem procesa terminalnih dodataka, pri čemu su Digenea preživjeli grabljivice svojih domaćina mehkušaca, vjerojatno pomoću riboe. Ostali domaćini dodani su istim postupkom sve dok se nije razvila moderna nevjerojatna raznolikost obrazaca životnog ciklusa.
Samo oko 12 od 6.000 poznatih vrsta su zarazne za ljude, ali neke od tih vrsta su važne za bolesti od kojih koja pati od 200 miliona ljudi. Vrste koje zaraze ljude mogu se podijeliti u skupine, shistosome i neshistosome.
Shistosomi se javljaju u istemu curkulacije konačnog domaćina. Ljudi se zaraze nakon što slobodnoplivjuća cerkarija oslobođenaiz zaraženih puževa prodre u kožu. Ovi dionoliki crvi su dugi i tanki, veličine su od 10 do 30 mm, a promjera od 0,2 do 1,0 mm. Mužjaci odraslih su kraći i deblji od ženki i imaju dugi utor duž jedne strane tijela u koji je ženka upletena. Ženke postižu spolnu zrelost nakon što se parenja. Nakon toga njih dvoje ostaju uvezani do kraja života. Mogu živjeti nekoliko godina i proizvesti mnogo hiljada jaja.
Četiri vrste shistosoma koji inficiraju ljude članovi su roda Schistosoma .
Shistosome Naučni naziv Prcvi međudomaćin Endemsko područje Schistosoma mansoni Biomphalaria spp. Afrika, Južna Amerika, Karibi, Srednji istok Schistosoma haematobium Bulinus spp. Afrika, Srednji istok Schistosoma japonicum Oncomelania spp. Kina, Istočna Azija, Filipini Schistosoma intercalatum Bulinus spp. AfrikaU nastavku je navedeno sedam glavnih vrsta neshistosoma koji inficiraju ljude. Ljudi se zaraze nakon gutanja metacerkarijskih cista na biljkama ili u pothlađenom životinjskom mesu. Većina vrsta nastanjuje ljudski gastrointestinalni trakt, gdje odlaže jaja koja u vanjsku sredinu odlaze zajedno s izmetom domaćina. "Paragonimus westermani", koji kolonizira pluća, također može odložiti jajašca u pljuvačku. Ove obično uzrokuju blage patološke posljedice kod ljudi, ali mogu se pojaviti i ozbiljniji učinci.
Neshistosome Naučni naziv Prvi međudomaćin Način infekcije Endemsko područje Fasciolopsis buski Segmentina sp. Biljke Azija, Indija Heterophyes heterophyes Pirinella{ Cipol, Tilapija Azija, Istočna Evropa, Egipat, Srednji istok Metagonimus yokogawaii Semisulcospira sp. Šaran, Pastrmke Sibir Gastrodiscoides hominis Helicorbis sp. Biljke Indija, Vijetnam, Filipini Clonorchis sinensis Bulinus sp. Ribe Istočna Azija, Sjeverna Amerika Fasciola hepatica Galba truncatula Biljke Širom svijeta Paragonimus westermani Oncomelania sp. Moski i slatkovodni rakovi AzijaDigenea uključuje oko 80 porodica.[3] Slijedi spisak po redovima: Digenea
Digenea (grč. dis – dvostruko, genos – pleme) je razred reda Trematoda u koljenu Platyhelminthes . Sastoji se od parazitskih pljosnatih glista s (poznatog kao metilji ) sa sincicijskim tegumentom (tjelesnim omotačem) i obično, dvije pijavke, jedna ventralna i jedna usna. Odrasli obično žive u probavnom traktu, ali se javljaju u svim organima svih razreda kičmenjaka. Nekada se mislilo da su povezane sa Monogenea, ali je prepoznato da su one najbliže grupi Aspidogastrea i da su monogenee bliže Cestoda. Do danas je opisano oko 6 000 vrsta.
Трематодтар (лат. Digenea, (рус. трематоды ) — Plathelminthes яҫы селәүсендәр синыфы (Plathelminthes), дегенетик имеүселәр. Уларҙың йәшәү циклы, һирәк осраҡты иҫәпкә алмағанда, бер нисә хужала кәм тегәндә өс быуын алышына. 7200-нән артыҡ төрө билдәле. Ҡайһы бер трематодтар трематодоз, шистосомоз , описторхоз кеүек ауырыу тыуҙырыусы кеше паразиттары. Кеше паразиттарының 40-тан артыҡ төрө бар.
Трематодтар (лат. Digenea, (рус. трематоды ) — Plathelminthes яҫы селәүсендәр синыфы (Plathelminthes), дегенетик имеүселәр. Уларҙың йәшәү циклы, һирәк осраҡты иҫәпкә алмағанда, бер нисә хужала кәм тегәндә өс быуын алышына. 7200-нән артыҡ төрө билдәле. Ҡайһы бер трематодтар трематодоз, шистосомоз , описторхоз кеүек ауырыу тыуҙырыусы кеше паразиттары. Кеше паразиттарының 40-тан артыҡ төрө бар.
De Digenea zijn een onderklasse van parasitaire platwormen (Platyhelminthes). Hiertoe behoort onder andere de familie Schistosomatidae waarin diverse soorten zijn die de veroorzakers zijn van de ziekte bilharzia.
Onder invloed van modern moleculair-genetisch onderzoek is de fylogenie van deze groep onderhevig aan grote veranderingen en is iedere keuze voor een indeling aanvechtbaar.
De Digenea zijn een onderklasse van parasitaire platwormen (Platyhelminthes). Hiertoe behoort onder andere de familie Schistosomatidae waarin diverse soorten zijn die de veroorzakers zijn van de ziekte bilharzia.
Przywry wnętrzniaki, Przywry digenetyczne (Digenea) – podgromada płazińców obejmująca bezwarunkowe pasożyty wewnętrzne kręgowców o ciele nieczłonowanym i listkowanym.
Mają z reguły dwie przyssawki stanowiące aparat czepny - okołogębową i brzuszną. Tylko u nielicznych przywr występuje jedna przyssawka[1] . Jelito zazwyczaj rozgałęzia się na dwa główne ramiona. Przywry pokryte są komórkami nabłonka nie posiadającymi rzęsek i zlewającymi się w jednolitą warstwę, czyli syncytium. Układ pokarmowy jest podobny do układu wirków choć bez wysuwanej gardzieli.
Niektóre gatunki przywr:
Charakteryzuje je złożony cykl rozwojowy z co najmniej dwoma żywicielami, przy czym jednym z nich jest zazwyczaj mięczak. Występują kolejno stadia miracidium, sporocysty, redii, cerkarii i metacerkarii.
Przywry wnętrzniaki, Przywry digenetyczne (Digenea) – podgromada płazińców obejmująca bezwarunkowe pasożyty wewnętrzne kręgowców o ciele nieczłonowanym i listkowanym.
Digenea Trematodaların (karaciğer kelebeklerinin) alt sınıfıdır. Bu alt sınıftaki parazitler erişkin memeli ve kanatlıların endoparazitlerindendir.Bu hayvanların karaciğer, sindirim kanalı ve dolaşım sisteminde yaşarlar. Larva dönemlerinden bazıları doğada serbest halde bazıları ise arakonakta bulunur.
Erişkinler: Dış görünüşleri genelde yaprak şeklinde olmakla beraber tesbih tanesi (örn: paramphistomum spp.) veya ince uzun silindirik (örn: shistosoma) olanları da vardır.
Vücut üzerinde tegüment adı verilen bir yapı bulunur. Bu yapının bazı görevleri vardır :
Vücutlarında ayrıca karın çekmeni, ağız çekmeni ve genital çekmen bulunabilir. Bu çekmenlerin görevi konağa tutunmayla ilgilidir.
Vücut boşlukları olmadığından organlar parankima içerisinde yerleşmiştir. Sindirim sistemleri basittir.Agız ile başlar, prefarenks, farenks, özafagus ve 2 kol halinde gelen bağırsaklar halinde devam eder. Anüs bulunmaz.Sindirim bağırsaklarda olur. Bu nedenle sindirilmemeş besinler ağızdan dışarı atılır. Sinir sistemleri uzunlamasına seyreden iki sinir ipliği ile bunların özafagus civarında birleşmesinden oluşan iki sinir yumağından ibarettir.
Schistosomoidea hariç hermafroditlerdir.
Digenea Trematodaların (karaciğer kelebeklerinin) alt sınıfıdır. Bu alt sınıftaki parazitler erişkin memeli ve kanatlıların endoparazitlerindendir.Bu hayvanların karaciğer, sindirim kanalı ve dolaşım sisteminde yaşarlar. Larva dönemlerinden bazıları doğada serbest halde bazıları ise arakonakta bulunur.
Дігенеї містять як мінімум 50 родин.
Acanthocolpidae Derogenidae Haplosplanchnidae Paramphistomidae Accacoeliidae Dicrocoeliidae Hemiuridae Paragonimidae Allocreadiidae Didymozoidae Heronomidae Philophthalmidae Angiodictyidae Diplostomidae Heterophyidae Plagiorchiidae Apocreadiidae Echinostomatidae Lecithasteridae Sanguinicolidae Atractotrematidae Enenteridae Lepocreadiidae Schistosomatidae Azygiidae Fasciolidae Microphallidae Sclerodistomidae Bivesiculidae Faustulidae Monorchiidae Strigeidae Bucephalidae Fellodstomidae Nasitrematidae Syncoeliidae Campulidae Gorgoderidae Notocotylidae Tandanicolidae Cryptogonimidae Gyliauchenidae Opecoelidae Transversotrematidae Cyclocoelidae Haploporidae Opisthorchiidae ZoogonidaeОкремі родини є базальними, або incertae sedis:
Дигенетические сосальщики[2] (лат. Digenea) — подкласс паразитических плоских червей из класса трематод (Trematoda)[1]. Их жизненный цикл, за редкими исключениями, протекает в нескольких хозяевах и сопровождается закономерным чередованием не менее трёх поколений. Описано около 7200 видов. Некоторые трематоды (около 40 видов) — паразиты человека, вызывающие опасные заболевания — трематодозы, например, шистосомоз и описторхоз. К трематодам относятся печёночная двуустка (Fasciola hepatica), кошачья двуустка (Opisthorchis felineus), шистосомы (Schistosoma), Leucochloridium paradoxum.
Раньше, особенно в русскоязычной научной традиции, название трематоды (Trematoda) использовалось как синоним дигенетических сосальщиков[3]. В современной гельминтологии этим термином называют класс плоских червей, объединяющий подклассы аспидогастрей (Aspidogastrea) и дигенетических сосальщиков (Digenea)[4].
К классу дигенетических сосальщиков относятся самые мелкие представители плоских червей, взрослая особь мелких видов может достигать 0,2 мм, крупных видов — 0,6 мм[5].
Дигенетические сосальщики обладают сложным жизненным циклом с гетерогонией: закономерным чередованием амфимиктной (обычно гермафродитной) и партеногенетических стадий. Всех представителей класса характеризует наличие стадий, паразитирующих в беспозвоночных (обычно в брюхоногих моллюсках). У большинства видов в жизненном цикле также присутствует, по меньшей мере, ещё один хозяин — позвоночное.
Из яйца выходит особь первого партеногенетического поколения — мирацидий, который способен плавать с помощью биения ресничек. Для дальнейшего развития мирацидий должен заразить следующего хозяина, в котором будут паразитировать все партеногенетические стадии. В роли этого хозяина практически у всех дигенетических сосальщиков выступают брюхоногие моллюски. Лишь некоторые группы в качестве хозяина используют двустворчатых моллюсков (например, из семейства Bucephalidae) или многощетинковых червей (из семейства Aporocotylidae). Эту стадию характеризует высокая специфичность к хозяину: как правило, мирацидии заражают и развиваются в моллюсках лишь одного вида (или нескольких близкородственных видах).
Выход мирацидия из яйца у разных видов происходит либо после заглатывания яйца хозяином (пассивное заражение), либо во внешней среде (активное заражение; подразумевает поиск хозяина). Последний вариант может реализоваться только у тех видов, яйца которых для развития должны попасть в воду. В то же время, заражение даже водных моллюсков некоторыми видами трематод осуществляется пассивно.
При обнаружении хозяина мирацидий проникает через покровы и претерпевает метаморфоз, обычно сводящийся к сбрасыванию ресничных покровов и формированию нового эпителия — тегумента. У небольшого числа видов превращение носит более кардинальный характер, и их мирацидии теряют почти все органы, а следующие стадии развиваются из небольшого числа клеток. Претерпевший метаморфоз мирацидий называют материнской спороцистой. В материнской спороцисте из партеногенетических яйцеклеток развивается следующее поколение — дочерние партениты, в зависимости от строения называемые редиями или дочерними спороцистами. Дочерние партениты в размножения отрождают либо следующие поколения дочерних партенит такого же строения, либо церкарий — личинок амфимиктного поколения.
Дочерние партениты некоторых трематод могут обитать в моллюске в течение нескольких лет, не приводя к его гибели, подавляя, однако, его репродуктивную активность. Паразитизм других видов, характеризующихся массовым выходом церкарий, закономерно приводит к гибели хозяина.
Поведение вышедшей из хозяина церкарии (расселительной личинки) специфично для разных видов. Выделяют три основных стратегии:
Существуют дигенетические сосальщики (например, лейкохлоридий парадоксальный), церкарии которых не выходят из тела хозяина, а заражают следующего хозяина при поедании им предыдущего.
За исключением единичных случаев, окончательный хозяин, в котором развиваются половозрелые особи амфимиктного поколения (мариты), — позвоночное. В организме хозяина они могут локализоваться в кишечнике, паренхиме и протоках печени, жёлчном пузыре, органах выделительной системы, дыхательных путях, кровеносном русле или даже под веком. Образующиеся после скрещивания марит яйца выводятся во внешнюю среду с калом или мочой хозяина.
В 2010 году американские исследователи показали, что партениты одного из видов рода Himasthla, паразитирующие в улитках Cerithidea californica, чётко разделяются на две группы, характеризующиеся различными размерами, строением и поведением. Более крупные редии ведут малоподвижный образ жизни и способны к размножению, более многочисленные мелкие редии, названные первооткрывателями «солдатами», не размножаются и активно атакуют как плоских червей других видов, так и неродственных им представителей того же вида[6].
Однако, данная концепция подвергается критике со стороны многих специалистов по гельминтам. В качестве основных аргументов, демонстрирующих несостоятельность вывода о наличии у редий разделения на настоящие касты, можно привести следующие наблюдения:
Дигенетические сосальщики (лат. Digenea) — подкласс паразитических плоских червей из класса трематод (Trematoda). Их жизненный цикл, за редкими исключениями, протекает в нескольких хозяевах и сопровождается закономерным чередованием не менее трёх поколений. Описано около 7200 видов. Некоторые трематоды (около 40 видов) — паразиты человека, вызывающие опасные заболевания — трематодозы, например, шистосомоз и описторхоз. К трематодам относятся печёночная двуустка (Fasciola hepatica), кошачья двуустка (Opisthorchis felineus), шистосомы (Schistosoma), Leucochloridium paradoxum.
Раньше, особенно в русскоязычной научной традиции, название трематоды (Trematoda) использовалось как синоним дигенетических сосальщиков. В современной гельминтологии этим термином называют класс плоских червей, объединяющий подклассы аспидогастрей (Aspidogastrea) и дигенетических сосальщиков (Digenea).
複殖亞綱(学名:Digenea)是扁形动物门吸蟲綱的一個亞綱,也是吸蟲綱絕大多數物種所屬。是一種寄生性扁蟲。截至目前,已包括有約六千個已紀錄的物種。
其物種泛稱複殖吸蟲(digenetic trematode),是主要以寄生人體為對象的吸蟲。複殖吸蟲的基本結構和發育過程大致上相同,其物種的雄性生殖系统的输精管的数目有兩個。本目物種種類相當繁多且形態各自相差頗大,生活史有性世代(寄生於軟體動物,如螺螄、蚌類的階段)與無性世代(寄生於脊椎動物體內階段),基本生活型態是以卵生且多種幼蟲(毛蚴、胞蚴、雷蚴、尾蚴、囊蚴)之後再成長為成蟲[1]。這些吸虫中以人类为宿主[2]的种类有[1]:肺吸虫(卫氏肺吸虫)、肝吸虫(中華肝吸蟲,牛羊肝吸蟲)、肠道吸虫、横川吸虫、血吸虫(日本血吸虫)、薑片蟲(Fasciolopsis buski)等。
Digenea (Gr. Dis – double, Genos – race)
複殖亞綱原為複殖目,又名二生目[2],當時分為兩個亞目。
現時之複殖亞綱下可分為五個目:
整個複殖亞綱現時包括有約80個科[3]。 以下為部分比較常見的科:
Acanthocolpidae Derogenidae Haplosplanchnidae 同盘科 ParamphistomatidaeThere are some digenean families that are either basal to or incertae sedis among the orders listed above:
對於目前已知的六千多種複殖亞綱物種來說,雖然當中只有12種會感染人類, 但當中有些是很嚴重的傳染病病源,影響人口高達二億人。這些可1感染人類的物種,大致可以分為以下兩類:
血吸蟲(Schistosomes)寄生於最終宿主的循環系統內。
會影響人類的七種非吸血吸蟲物種詳列如下。一般人都透過不慎進食了含有蟲卵的植物或未煮至熟透的動物肉類而感染。大多數物種寄居在人體的腸道內,並且在宿主的糞便裡產卵;衛氏肺吸蟲(Paragonimus westermani)雖然在人體的肺部繁殖,但其卵可透過飛沫傳播。這些物種儘管從病理學的角度來看會對人類構成沒那麼嚴重的問題,但亦可能會引起其他嚴重病變。
人類非血吸蟲 中文名 學名 第一宿主 怠染人類模式 影響地區 布氏薑片蟲 Fasciolopsis buski Segmentina(英语:Segmentina)屬物種 植物 亚洲(包括印度) Heterophyes heterophyes(英语:Heterophyes heterophyes) Pirinella[來源請求] 鯔形目物種、罗非鱼 亚洲、中东、埃及與东欧 Metagonimus yokogawaii Semisulcospira(英语:Semisulcospira)屬物種[來源請求] 鯉科、鮭科物種 西伯利亞 拟人腹盘吸虫 Gastrodiscoides hominis Helicorbis sp.[來源請求] 植物 印度、越南及菲律賓 華支睾吸蟲 Clonorchis sinensis 沼螺屬物種 魚類 東亞及北美 牛羊肝吸蟲 Fasciola hepatica 截口土蜗(Galba truncatula) 植物 全世界 衛氏肺吸蟲 Paragonimus westermani 釘螺屬物種 螃蟹及螫蝦 亞洲複殖亞綱(学名:Digenea)是扁形动物门吸蟲綱的一個亞綱,也是吸蟲綱絕大多數物種所屬。是一種寄生性扁蟲。截至目前,已包括有約六千個已紀錄的物種。
其物種泛稱複殖吸蟲(digenetic trematode),是主要以寄生人體為對象的吸蟲。複殖吸蟲的基本結構和發育過程大致上相同,其物種的雄性生殖系统的输精管的数目有兩個。本目物種種類相當繁多且形態各自相差頗大,生活史有性世代(寄生於軟體動物,如螺螄、蚌類的階段)與無性世代(寄生於脊椎動物體內階段),基本生活型態是以卵生且多種幼蟲(毛蚴、胞蚴、雷蚴、尾蚴、囊蚴)之後再成長為成蟲。這些吸虫中以人类为宿主的种类有:肺吸虫(卫氏肺吸虫)、肝吸虫(中華肝吸蟲,牛羊肝吸蟲)、肠道吸虫、横川吸虫、血吸虫(日本血吸虫)、薑片蟲(Fasciolopsis buski)等。




