Baculoviridae é unha familiade virus comunmente chamados baculovirus, que teñen como hóspedes naturais artrópodos, como insectos lepidópteros, himenópteros, dípteros e crustáceos decápodos. Actualmente existen nesta familia 66 especies, repartidas en 4 xéneros.[1][2]
Os baculovirus infectan invertebrados, e describíronse unhas 600 especies de hóspedes. As formas inmaduras (larvarias) de especies de avelaíñas son os hóspedes máis comúns, pero estes virus tamén infectan sínfitos, mosquitos, e gambas. Aínda que os baculovirus poden entrar en células de mamíferos en cultivo[3] non poden replicarse en células de vertebrados.
A partir da década de 1940 foron estudados e utilizados como biopesticidas en agricultura. Os baculovirus conteñen un xenoma de ADN bicatenario de entre 80 e 180 kbp.
As mencións máis antigas da probable presenza de baculovirus poden encontrarse xa na literatura desde inicios do século XVII en informes dunha enfermidade que afectaba os vermes da seda. A partir da década de 1940 foron moi usados e estudados como biopesticidas nos campos agrícolas. A partir da década de 1990 foron utilizados para producir proteínas eucariotas complexas en cultivos celulares de células de insectos (ver Sf21, Hi-5). Estas proteínas recombinantes foron utilizadas en investigación como vacinas tanto en humanos coma en tratamentos veterinarios (por exemplo, a vacina máis amplamente utilizada para a prevención da gripe aviaria por H5N1 en polos foi producida nun vector de expresión baculovirus). Máis recentemente atopouse que os baculovirus poden transducir células de mamíferos cun promotor axeitado.[4] Estes usos médicos aplicados ou potenciais fixeron que aumentase o número de publicacións sobre baculovirus desde 1995.
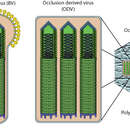
O ciclo de vida dos baculovirus implica dúas formas distitnas do virus. O virus derivado da oclusión (ODV) está presente nunha matriz proteica (poliedrina ou granulina) e é responsable da infección primaria do hóspede, mentres que o virus evaxinado (BV) libérase das células hóspede infectadas posteriormente durante a infección secundaria.
Os baculovirus teñen tropismos moi específicos de especie entre os invertebrados e describíronse unhas 700 especies hóspede. As forma inmaduras (larvas) das avelaíñas son os hóspedes máis comúns, pero estes virus infectan tamén a sínfitos, mosquitos e gambas.
Tipicamente, a infección inicial ocorre cando un insecto hóspede susceptible se alimenta de plantas que están contaminadas coa forma ocluída do virus. A matriz proteica disólvese no ambiente alcalino do estómago do hóspede, liberando ODVs que despois se fusionan coas membranas das células epiteliais columnares do intestino do hóspede e penetran na célula dentro de endosomas. As nucleocápsides escapan dos endosomas e son transportadas ao núcleo. Este paso está mediado posiblemente por filamentos de actina. A transcrición viral e a replicación ocorren no núcleo celular e evaxínanse novas partículas de BV desde a membrana basolateral para espallar a infección sistematicamente. Durante a evaxinación, os BV adquiren unha membrana da célula hóspede que está axustada de forma frouxa e contén glicoproteínas virais.
A infección por baculovirus pode ser dividida en tres fases:
Aínda que os BV se producen na fase tardía, a forma ODV prodúcese na fase moi tardía, na que adquire a envoltura do núcleo da célula hóspede e é incrustada na matriz da proteína do corpo de inclusión. Estes corpos de inclusión son liberados cando as células se lisan para espallar a infección por baculovirus aos seguintes hóspedes. A extensa lise de células causa frecuentemente que o insecto hóspede literalmente se desintegre. As partículas completas de ODV-poliedrina son resistentes á inactivación pola calor e a luz, mentres que o virión BV espido é máis sensible ás condicións do ambiente.
Cando infectan unha eiruga, os estadios avanazados da infección causan que o hóspede se alimente sen parar e despois gabee ás partes máis altas das árbores, includíndo sitios expostos que normalmente evitan debido ao risco que representan os predadores. Esta é unha vantaxe para o virus se (cando o hóspede se disolve) este pinga sobre as follas de máis abaixo que son consumidas por novos hóspedes.[5]
O baculovirus máis estudado é o Nucleopoliedrovirus multicápside de Autographa californica (AcMNPV, Autographa californica multicapsid nucleopolyhedrovirus). O virus foi illado orixinalmente do lepidóptero Autographa californica e contén un xenoma de 134-kbp con 154 marcos abertos de lectura (ORF). A principal proteína da cápside é a VP39, que xunto con algunhas proteínas menores forma a nucleocápside (21 nm x 260 nm) que encerra o ADN coa proteína p6.9.
O BV adquire a súa envoltura da membrana da célula hóspede e necesita a glicoproteína gp64 para poder espallarse e xerar unha infección sistémica. Esta proteína forma estruturas chamadas peplómeros nun dos extremos da partícula do virus evaxinado pero non se encontra no ODV (aínda que outras proteínas están só asociadas coa forma ODV). Existen tamén algunhas diferenzas na composición lipídica da envoltura viral das dúas formas. Aínda que a envoltura do BV consta de fosfatidilserina, o ODV contén fosfatidilcolina e fosfatidiletanolamina.
Identificouse un elemento esencial para a ensamblaxe da nucleocápside (NAE) no xenoma da AcMNPV. O NAE é un elemento en cis interno dentro do xene ac83. A ensamblaxe da nucleocápside non depende da produción da proteína Ac83.[6]
Durante a evolución, as glicoproteínas da envoltura baculoviral non sufriron cambios. A Ld130, tamén chamada proteína F de baculovirus de Lymantria dispar (LdMNPV) suxeriuse que é unha proteína de fusión de envoltura ancestral que foi substituída por medio dunha substitución de xene non ortólogo por gp64 no AcMNPV, Bombyx mori (BmNPV) e Orgyia pseudotsugata (OpMNPV), mentres que aínda reteñen o xene ld130.
A Gp64 é unha glicoproteína de membrana homotrimérica, que está polarmente presente sobre o virión con forma de bastón. Consta de 512 aminoácidos (aa) con catro sitios de glicosilación en residuos de asparaxina e ten unha secuencia sinal N-terminal de (20 aa), a oligomerización e o dominio de fusión e un dominio transmembrana hidrófobo preto do C-terminal (7 aa).
É producido tanto nas fases temperá coma na tardía do ciclo de infección e chega á velocidade máxima de síntese entre as 24 e 26 horas post-infección. A trimerización con enlaces disulfuro de cisteína intermoleculares parece ser un paso crucial para o transporte da proteína á superficie celular, dado que só o 33% da proteína sintetizada chega á superficie celular, xa que a gp64 monomérica é degradada dentro das células.
A gp64 é esencial para a evaxinación eficiente do virión e para a transmisión de célula a célula durante o ciclo de infección e para a entrada viral, é dicir, causando un trofismo viral e a captación pola célula mediada por endosomas. A función principal da proteína da envoltura gp64 é causar a fusión mediada polo pH da envoltura e o endosoma. Aínda que o gp64 ten varias funcións esenciais, informouse que nos baculovirus nulos para a gp64 poden substituírses esta por outras glicoproteínas virais como a Ld130, a proteína G do virus da estomatite vesicular. Estas substitucións teñen como resultado a formación de virións funcionais.
A expresión dos baculovirus en células de insectos representa un método robusto para producir glicoproteínas recombinantes ou proteínas de membrana.[7][8][9] As proteinas producidas por baculovirus están a ser estudados actualmente como vacinas terapéuticas contra o cancro con varias vantaxes inmunolóxicas sobre as proteínas derivadas de fontes de mamíferos.[10]
Os baculovirus non poden replicarse dentro de células de mamíferos ou plantas.[11] Teñen un rango de hóspedes restrinxido aos que poden infectar normalmenre limitado a un número limitado de especies de insectos estreitamente relacionadas. Como os baculoviruses non son nocivos para os humanos, son considerados unha opción segura para o seu uso nas aplicacións de investigación. Tamén se usan como axentes biolóxicos como no caso da loita contra a pirálida Plodia interpunctella, unha praga dos cereais.[12]
O nome desta familia deriva do latín baculus, que significa bastón. A familia foi dividida en catro xéneros: Alphabaculovirus (nucleopoliedrovirus específicos de lepidópteros), Betabaculovirus (granulovirus específicos de lepidópteros), Gammabaculovirus (nucleopoliedrovirus específicos de himenópteros) e Deltabaculovirus (nucleopoliedrovirus específicos de dípteros).[13] Ver a seguinte lista despregable:
Grupo: dsDNA (virus de ADN bicatenario)
Os baculovirus crese que evolucionaron a partir de virus da familia dos nudivirus (Nudiviridae) hai 310 millóns de anos.[15]
Baculoviridae é unha familiade virus comunmente chamados baculovirus, que teñen como hóspedes naturais artrópodos, como insectos lepidópteros, himenópteros, dípteros e crustáceos decápodos. Actualmente existen nesta familia 66 especies, repartidas en 4 xéneros.
Os baculovirus infectan invertebrados, e describíronse unhas 600 especies de hóspedes. As formas inmaduras (larvarias) de especies de avelaíñas son os hóspedes máis comúns, pero estes virus tamén infectan sínfitos, mosquitos, e gambas. Aínda que os baculovirus poden entrar en células de mamíferos en cultivo non poden replicarse en células de vertebrados.
A partir da década de 1940 foron estudados e utilizados como biopesticidas en agricultura. Os baculovirus conteñen un xenoma de ADN bicatenario de entre 80 e 180 kbp.