Ecology
provided by NMNH Marine Dinoflagellates
D. acuminata is a planktonic toxic bloom-forming species (Taylor et al. 1995; Steidinger & Tangen 1996). The most extensive blooms have been reported from the summer and fall months (Kat 1989; Taylor et al. 1995). Blooms have been reported from many parts of the world (see Kat 1985); however, they have been particularly extensive with cell concentrations less than 40,000 cells/L (Kat 1985; 1989). Blooms are often associated with toxicity of shellfish (Taylor et al. 1995). Jacobson and Andersen (1994) found a high number of food vacuoles in cells of Dinophysis acuminata and deduced that mixotrophy is an important aspect of its biology. They speculate that this species feeds by way of a peduncle (myzocytosis), the feeding mode used by the heterotrophic species Dinophysis rotundata and D. hastata (Schnepf & DeichgrAbèr 1983). The peduncle, the proposed feeding apparatus, passes through the cytostomal opening in the theca when the cell is feeding (Jacobson & Andersen 1994).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Habitat and Locality
provided by NMNH Marine Dinoflagellates
Populations of Dinophysis acuminata are distributed widely in temperate waters. They are most common and abundant in coastal waters of the northern Atlantic and Pacific Oceans, especially eutrophic areas (Taylor et al. 1995; Steidinger & Tangen 1996).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Morphology and Structure
provided by NMNH Marine Dinoflagellates
Dinophysis acuminata is a photosynthetic species with large chloroplasts, a posterior pyrenoid, and a large central nucleus (Hallegraeff & Lucas 1988; Zingone et al. 1998).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Nomenclatural Types
provided by NMNH Marine Dinoflagellates
Holotype: Dinophysis acuminata Claparède and Lachmann, 1859: 408, plate 20, fig. 17
Type Locality: North Sea: Norway
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Remarks
provided by NMNH Marine Dinoflagellates
D. acuminata has a history wrought with identification problems mainly attributable to the morphological variability of this species. This problem is enhanced by the many synonyms and questionable identifications that have accumulated in the literature over the years (see Zingone et al. 1998). Compounding the identification problem is the influence of feeding on lateral cell shape; cells containing food vacuoles had a rounder lateral outline than cells devoid of food vacuoles (Jacobson & Andersen 1994). Many authors consider Phalacroma to be synonymous with Dinophysis (Steidinger & Tangen 1996).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Reproduction
provided by NMNH Marine Dinoflagellates
D. acuminata reproduces asexually by binary fission. Mackenzie (1991) reported sexual reproduction via the fusion of anisogamous gametes.
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Species Comparison
provided by NMNH Marine Dinoflagellates
D. acuminata can be confused with D. sacculus, D. norvegica, D. ovum and D. punctata, but is most often misidentified as D. sacculus (Steidinger & Tangen 1996; Zingone et al. 1998). The major difference between D. acuminata and D. sacculus is the shape of the large hypothecal plates: in D. acuminata they are shorter, more convex dorsally and often more slender posteriorly; whereas, in D. sacculus they are long and sack-like. D. acuminata also exhibits more pronounced thecal areolation and sulcal list ornamentation, but these are variable features. Since these two species rarely occur in the same area with the same importance, the possibility of misidentification is reduced (Zingone et al. 1998). Surface thecal ornamentation in this species is similar to D. sacculus (Hallegraeff & Lucas 1988).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Species Overview
provided by NMNH Marine Dinoflagellates
Dinophysis acuminata is an armoured, marine, planktonic dinoflagellate species. It is a toxic species associated with DSP events and is commonly found in coastal waters of the northern Atlantic and Pacific Oceans.
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Synonyms
provided by NMNH Marine Dinoflagellates
Dinophysis lachmannii Paulsen, 1949
Dinophysis borealis Paulsen, 1949
Dinophysis boehmii Paulsen, 1949
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Taxonomic Description
provided by NMNH Marine Dinoflagellates
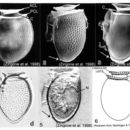
Species in this genus are laterally compressed with a small, cap-like epitheca and a much larger hypotheca (dorso-ventral depth of epitheca is 1/3 to 1/2 of hypotheca). The shape of the cell in lateral view is the most important criterion used for identification (Taylor et al. 1995). However, size and shape varies considerably in this species (Larsen & Moestrup 1992).
Cells of Dinophysis acuminata are small to medium, almost oval or elliptical in shape (Figs. 1-5). The shape can vary from rotund to long and narrow in lateral view. A well-developed left sulcal list (LSL) extends beyond the midpoint of the cell (1/2 to 2/3 of cell length) (Figs. 1-3). The antapex is rounded, and cells are commonly found with two to four small knob-shaped posterior protrusions; sometimes well-developed and sometimes not (Figs. 2-5) (Balech 1976; Hallegraeff & Lucas 1988; Taylor et al. 1995; Steidinger & Tangen 1996).
The thick thecal plates are covered with prominent circular areolae, each with a pore (Fig. 2). These markings can vary depending on the age of the cell. The variations can range from only pores (Fig. 3), to depressions with scattered pores (Fig. 1), to depressions each with a pore, to areolae each with a pore (Fig. 2). Pores are not found in the megacytic zone (Fig. 3). Cell size ranges: 38-58 µm in length and 30-40 µm in dorso-ventral width (widest near middle of cell) (Lebour 1925; Abè 1967; Dodge 1982; Fukuyo et al. 1990; Larsen & Moestrup 1992; Taylor et al. 1995; Steidinger & Tangen 1996).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Thecal Plate Description
provided by NMNH Marine Dinoflagellates
The epitheca is slightly convex and inclined ventrally (Figs. 1-4). Made up of four plates, it is not visible in lateral view (Balech 1976; Hallegraeff & Lucas 1988; Taylor et al. 1995; Zingone et al. 1998).
The cingulum is made up of four unequal plates, and is bordered by two well-developed lists: an anterior cingular list (ACL), often with ridges, and a smooth posterior cingular list (PCL) (Fig. 1). The dorsal end of the cingulum is concave, strongly inclined and (Figs. 1, 6) (Balech 1976; Zingone et al. 1998).
The sulcus is comprised of four irregularly shaped plates. The flagellar pore is housed in the sulcal area. The LSL, supported by three ribs, is rather narrow and often sculptured with reticulated ribs, lines and areolae (Balech 1976; Taylor et al. 1995; Zingone et al. 1998). The third rib on the left sulcal list is the longest, and is usually strongly curved posteriorly (Figs. 1, 4, 6). Sulcal plate development is highly variable in this species (Balech 1976).
The hypotheca, with four large plates, comprises the majority of the cell. The dorsal margin is more or less evenly convex (Figs. 1, 2, 4). The ventral margin is rarely convex; it is generally oblique and flat (Figs. 2-5)(Balech 1976). The antapex is ventrally off-center (Figs. 2-5)(Abè 1967).
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Toxicity
provided by NMNH Marine Dinoflagellates
D. acuminata is a toxic species that has been found to produce okadaic acid (OA) (Cembella 1989; Lee et al. 1989) causing diarrhetic shellfish poisoning (DSP) (Kat 1985). Toxicity can vary considerably among seasons and areas where it blooms (Taylor et al. 1995). This species can cause shellfish toxicity at very low cell concentrations (as low as 200 cells/L)(Lassus et al. 1985). Hoshiai et al. (1997), however, reported a case of nontoxic mussels in Kesennuma Bay, northern Japan, in the presence of high concentrations of D. acuminata cells.
- bibliographic citation
- Faust, Maria A. and Rose A. Gulledge. Identifying Harmful Marine Dinoflagellates. Smithsonian Contributions from the United States National Herbarium, volume 42: 1-144 (including 48 plates, 1 figure and 1 table).
Dinophysis acuminata
provided by wikipedia EN
Dinophysis acuminata is a marine plankton species of dinoflagellates that is found in coastal waters of the north Atlantic and Pacific oceans.[1] The genus Dinophysis includes both phototrophic and heterotrophic species. D. acuminata is one of several phototrophic species of Dinophysis classed as toxic, as they produce okadaic acid which can cause diarrhetic shellfish poisoning (DSP). Okadiac acid is taken up by shellfish and has been found in the soft tissue of mussels and the liver of flounder species. When contaminated animals are consumed, they cause severe diarrhoea. D. acuminata blooms are constant threat to and indication of diarrhoeatic shellfish poisoning outbreaks.[2][3][4]
Dinophysis acuminata is a photosynthesising Dinophysis species by acquiring secondary plastids from consuming the ciliate Myrionecta rubra,[5] which in turn had ingested them from the alga Teleaulax amphioxeia.[6][7] Thus, D. acuminata is a mixotroph, primarily a heterotroph, but autotroph once it acquires plastids. This is also an example of cell organelle stealing, the concept called kleptoplasty, and endosymbiosis. Dinophysis acuminata reproduces sexually and asexually.[8]
Description
Dinophysis acuminata is an oval-shaped protist. It measures 30-35 μm in length and 38-58 μm in diameter. The body is reddish-brown in colour and is covered with an armor-like covering called theca, which is made up of grass. The anterior end has a crown-like platform, which is the smaller epitheca; while the posterior is simply rounded constituting a larger hypotheca. The cell has two flagella for locomotion. Reproduction is by simple binary fission. In lateral view D. acuminata cells are irregularly egg-shaped, dorsally convex and have large hypothecal plates with a more or less oval shape. The dorsal contour is always more strongly convex than the ventral one. Compared to other species of Dinophysis, D. acuminata has a more straight ventral margin and larger left sulcal lists with more prominent ribs. The nucleus is prominently at the centre of the cell. The unusual feature of the cell is that it contains reddish-brown chloroplast.[1]
The taxonomic identification of Dinophysis species is largely based on cell contouring, size and shape of their large hypothecal plates and the shape of their left sulcal lists and ribs. When viewed laterally species in the Dinophysis are laterally compressed with a cap-like epitheca and a much larger hypotheca although the size and shape of these species varies greatly due to their polymorphic life cycle. Due to the morphological variability of Dinophysis species identification can be hard, especially when two species (D. acuminata and D. sacculus) co-exist. For this reason the term "D. acuminata complex" was coined to label a group of co-existing species difficult to discriminate.[9]
Dinophysis morphology
Dinophysis acuminata can be very hard to identify, and requires careful observations[3]. It can be identified by its midsection[1]. It is very large (38-55μm) and wide (30-38μm) in the middle unlike D. norvegica that is 34-50μm long and 36-43μm wide[2]. The Dinophysis norvegica is smaller and widest in the middle region[1]. D. norvegica and D. acuminata are a very similar species as a result, they exhibits similar behaviors and are commonly misidentified[1]. Furthermore, other ways to identify D. acuminata from other Dinophysis species can be done by comparison of the left sulcal list (cellulose extensions of the cell[5]) and LSL identification in ribs[1].Cells have convex dorsal margins and small oval shaped cells and their thecal plates are covered with areolae (circular depression on the cellulose wall of a dinoflagellate[5]) each with a pore[4]. Continuously, the knob-shaped protrusions and round antapex (botton end of a dinoflagellate[5]) are ways to identify D.acuminata[4].
Feeding and endosymbiosis
Dinophysis acuminata is basically a heterotroph feeding on the ciliate Mesodinium rubrum. M. rubrum in turn feeds on green algae that contain plastids. (The endosymbiont is used by the ciliate for its own photosynthesis.)[10] Microscopic observations of live cells using established cultures revealed that D. acuminata uses a peduncle, extending from the flagellar pore, to extract the cell contents of the marine ciliate M. rubrum. After about 1 minute the trapped M. rubrum becomes immobile after which the D. acuminata slowly consumes the ciliate, over 1–2 hours, filling its vacuoles with the ciliate's cytoplasm.[9] The algal plastids are not destroyed by D. acuminata but use it for its own photosynthesis, thereby becoming an autotroph. However, unlike its prey M. rubum, it is not clear whether D. acuminata uses the plastids permanently or temporarily.[11][12] Food vacuoles found in the vacuoles of this primitive genus indicates that organisms in this genus are mixotrophs especially D. norvegica[1]. Mixotrophy is the ability of an organism to use different sources of carbon and energy instead of having a single mode of feeding (autotroph or heterotroph). However, certain species related to Dinophysis acuminata prefer one mode of feeding over another. Dinophysis rotundata uses myzocytosis to feed [1]. Their mixotrophic conditions and size are influenced by prey populations and hydrographic conditions [2].
Ecology and current threats of Dinophysis
Dinophysis acuminata has caused several problems in oceanic ecosystems. The main cause of DSP, diarrhetic shellfish poisoning, outbreaks in waters along Europe has been due to this species[3]. Likewise, the close knit sisters of Dinophysis acuminata called Dinophysis norvegica, a photosynthetic organism with yellow chloroplast and posterior nucleus, Dinophysis acuta, and Dinophysis fortii have also been known to cause the same problems as D. acuminata[1]. Dinophysis norvegica is a marine plankton dinoflagellate that is found in neritic waters[1]. This species of the Dinophysis genus is a bloom-forming toxic species[1]. Both species reproduce asexually by binary fission to make identical copies of itself[1]. Speculations of sexual dimorphism that is the difference between the female and male counterparts have allowed researchers to draw conclusion that species of this genus can undergo sexual reproduction[1]. The first record of DSP with Dinophysis acuminata and Dinophysis fortii was in 1980 in the Patagonian coast[4]. A year after the report, another occurrence of DSP unleashed in December 1993 and November 1994[4]. Dinophysis acuminata releases lipophilic shellfish toxins (LSTs) and have been found to cause trouble to ecological marines and aquaculture farmers[2]'. The lipophilic toxins accumulate in shellfishes and causes diarrhea and shellfish poisoning to consumers. Dinophysis acuminata is the cause of DSP in Brazil and creates a disturbing impact due to its long and early blooming species[4]. However, their presence is restricted around the spring and summer[4], but in higher concentrations in December. DSP is a particular kind food poisoning that causes severe gastrointestinal illness in humans and this is related to the ingestion of toxin contaminated shellfishes from contaminated water[4]. Some of the symptoms of DSP include diarrhea, stomach pain, vomiting, nausea and fever; reported human ingestion shows that the toxins are capable of causing stomach tumors and chronic problems to consumers[4]. Government involvement as a result of high concentrations of toxins in the Dinophysis toxic shellfish epidemic has caused economic crisis in Europe and the aquaculture industry'[2]'.
References
-
^ a b Setälä, Outi; Autio, Riitta; Kuosa, Harri; Rintala, Janne; Ylöstalo, Pasi (2005). "Survival and photosynthetic activity of different Dinophysis acuminata populations in the northern Baltic Sea". Harmful Algae. 4 (2): 337–350. doi:10.1016/j.hal.2004.06.017. ISSN 1568-9883.
-
^ Díaz, Patricio; Reguera, Beatriz; Ruiz-Villarreal, Manuel; Pazos, Yolanda; Velo-Suárez, Lourdes; Berger, Henrick; Sourisseau, Marc (2013). "Climate variability and oceanographic settings associated with interannual variability in the initiation of Dinophysis acuminata blooms". Marine Drugs. 11 (8): 2964–2981. doi:10.3390/md11082964. PMC 3766876. PMID 23959151.
-
^ Lee, Ka Jeong; Mok, Jong Soo; Song, Ki Cheol; Yu, Hongsik; Jung, Jee Hyung; Kim, Ji Hoe (2011). "Geographical and annual variation in lipophilic shellfish toxins from oysters and mussels along the south coast of Korea". Journal of Food Protection. 74 (12): 2127–2133. doi:10.4315/0362-028X.JFP-11-148. PMID 22186054.
-
^ Naustvoll, L.-J.; Gustad, E.; Dahl, E. (2012). "Monitoring of Dinophysis species and diarrhetic shellfish toxins in Flødevigen Bay, Norway: inter-annual variability over a 25-year time-series". Food Additives & Contaminants: Part A. 29 (10): 1605–1615. doi:10.1080/19440049.2012.714908. PMID 22891979.
-
^ Johnson, Matthew D.; Oldach, David; Delwiche, Charles F.; Stoecker, Diane K. (2007). "Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra". Nature. 445 (7126): 426–428. doi:10.1038/nature05496. PMID 17251979.
-
^ Janson, Sven (2004). "Molecular evidence that plastids in the toxin-producing dinoflagellate genus Dinophysis originate from the free-living cryptophyte Teleaulax amphioxeia". Environmental Microbiology. 6 (10): 1102–1106. doi:10.1111/j.1462-2920.2004.00646.x. PMID 15344936.
-
^ Nishitani, G.; Nagai, S.; Baba, K.; Kiyokawa, S.; Kosaka, Y.; Miyamura, K.; Nishikawa, T.; Sakurada, K.; Shinada, A.; Kamiyama, T. (2010). "High-level congruence of Myrionecta rubra prey and Dinophysis species plastid identities as revealed by genetic analyses of isolates from Japanese coastal waters". Applied and Environmental Microbiology. 76 (9): 2791–2798. doi:10.1128/AEM.02566-09. PMC 2863437. PMID 20305031.
-
^ "WoRMS - World Register of Marine Species - Dinophysis acuminata Claparède & Lachmann, 1859". www.marinespecies.org. Retrieved 2016-09-28.
-
^ a b Raho, Nicolás; Pizarro, Gemita; Escalera, Laura; Reguera, Beatriz; Marín, Irma (2008). "Morphology, toxin composition and molecular analysis of Dinophysis ovum Schütt, a dinoflagellate of the "Dinophysis acuminata complex"". Harmful Algae. 7 (6): 839–848. doi:10.1016/j.hal.2008.04.006. ISSN 1568-9883.
-
^ Dorrell, R. G.; Howe, C. J. (2012). "What makes a chloroplast? Reconstructing the establishment of photosynthetic symbioses". Journal of Cell Science. 125 (8): 1865–1875. doi:10.1242/jcs.102285. PMID 22547565.
-
^ Takishita, K; Koike, K; Maruyama, T; Ogata, T (2002). "Molecular evidence for plastid robbery (Kleptoplastidy) in Dinophysis, a dinoflagellate causing diarrhetic shellfish poisoning". Protist. 153 (3): 293–302. doi:10.1078/1434-4610-00106. PMID 12389818.
-
^ Wisecaver, Jennifer H; Hackett, Jeremiah D (2010). "Transcriptome analysis reveals nuclear-encoded proteins for the maintenance of temporary plastids in the dinoflagellate Dinophysis acuminata". BMC Genomics. 11 (1): 366. doi:10.1186/1471-2164-11-366. PMC 3017763. PMID 20537123.

- license
- cc-by-sa-3.0
- copyright
- Wikipedia authors and editors
Dinophysis acuminata: Brief Summary
provided by wikipedia EN
Dinophysis acuminata is a marine plankton species of dinoflagellates that is found in coastal waters of the north Atlantic and Pacific oceans. The genus Dinophysis includes both phototrophic and heterotrophic species. D. acuminata is one of several phototrophic species of Dinophysis classed as toxic, as they produce okadaic acid which can cause diarrhetic shellfish poisoning (DSP). Okadiac acid is taken up by shellfish and has been found in the soft tissue of mussels and the liver of flounder species. When contaminated animals are consumed, they cause severe diarrhoea. D. acuminata blooms are constant threat to and indication of diarrhoeatic shellfish poisoning outbreaks.
Dinophysis acuminata is a photosynthesising Dinophysis species by acquiring secondary plastids from consuming the ciliate Myrionecta rubra, which in turn had ingested them from the alga Teleaulax amphioxeia. Thus, D. acuminata is a mixotroph, primarily a heterotroph, but autotroph once it acquires plastids. This is also an example of cell organelle stealing, the concept called kleptoplasty, and endosymbiosis. Dinophysis acuminata reproduces sexually and asexually.
- license
- cc-by-sa-3.0
- copyright
- Wikipedia authors and editors