Die Canacidae sind eine Familie der Zweiflügler (Diptera) und gehören hier zu den Fliegen (Brachycera). Weltweit sind etwa 100 Arten dieser Gruppe bekannt, in Mitteleuropa ist ausschließlich die Art Xanthocanace ranula zu finden.
Die Vertreter von Xanthocanace ranula sind etwa 2,5 Millimeter lang und leben an der Meeresküste. Die Larven der Fliegen weiden hier den Algenbewuchs ab, während sich die Imagines fliegend über Salzwassertümpeln aufhalten. Sie fischen mit ihrem speziell dafür ausgestatteten Rüssel mit einem Reusenapparat Kleinstlebewesen von der Wasseroberfläche.
Die Canacidae sind eine Familie der Zweiflügler (Diptera) und gehören hier zu den Fliegen (Brachycera). Weltweit sind etwa 100 Arten dieser Gruppe bekannt, in Mitteleuropa ist ausschließlich die Art Xanthocanace ranula zu finden.
Die Vertreter von Xanthocanace ranula sind etwa 2,5 Millimeter lang und leben an der Meeresküste. Die Larven der Fliegen weiden hier den Algenbewuchs ab, während sich die Imagines fliegend über Salzwassertümpeln aufhalten. Sie fischen mit ihrem speziell dafür ausgestatteten Rüssel mit einem Reusenapparat Kleinstlebewesen von der Wasseroberfläche.
Canacidae, incorrectly Canaceidae, or beach flies, surf or surge flies, is a family of Diptera. There are 113 species in 12 genera.[3][4][5][6] The family now includes Tethininae as a subfamily.
For terms see Morphology of Diptera.
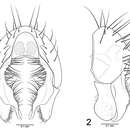
Minute (1.6–5 mm) yellow, grey or grey-brown pruinose flies with whitish to greyish markings. The head is large with small antenna bearing bare to pubescent arista. The "mouth" is a large oval opening. There are three or four pairs of orbital bristles on the head directed outward (inset upswept). Postvertical bristles are absent but diverging pseudopostocellar bristles are present. Other head bristles present are ocellar bristles, 2-5 pairs of frontal bristles, curving outward, interfrontal bristles and vibrissae ("whiskers"). The genae are high with 1 or more upcurving bristles. Tibiae are without a dorsal preapical bristle.
The wing is unmarked in almost all species. The costa has a subcostal break; the subcosta is parallel to vein R1 and merging with that vein just before the costa. Tibiae without dorsal preapical bristle.
See [1] Drawings of Canace.
Canacidae are mostly intertidal flies. They are found along sea coasts, on the surface of small water bodies, saline and fresh, at places protected from wind. They feed on Infusoria and other minute organisms.
Canacidae, incorrectly Canaceidae, or beach flies, surf or surge flies, is a family of Diptera. There are 113 species in 12 genera. The family now includes Tethininae as a subfamily.
 face on view of the head of Canace ranula, Loew
face on view of the head of Canace ranula, Loew  Wing venation
Wing venation
Les Canaceidae sont une famille de diptères.
Selon World Register of Marine Species (14 mars 2013)[2] :
Selon ITIS (14 mars 2013)[3] :
Canacidae Jones, 1906, è una famiglia di insetti dell'ordine dei Ditteri (Brachycera: Cyclorrhapha: Acalyptratae), simili nell'aspetto ai moscerini della frutta.
La maggior parte di questi insetti è associata ad ambienti costieri marini ed oceanici, perciò sono noti, in inglese, con nomi che fanno riferimento al loro tipico habitat, come beach flies, surf flies e surge flies ("mosche della spiaggia", "dell'onda", "della marea").
L'inquadramento sistematico di questa famiglia ha subito una recente revisione tassonomica con l'unione dei Canacidae sensu stricto e dei Tethinidae. Fino al 2007, infatti, i Canacidae sensu lato erano distinti in due famiglie affini, Canacidae sensu stricto e Tethinidae, ma già dagli anni ottanta si ipotizzava la possibile unione in un unico taxon. L'ipotesi di unione si è rafforzata negli anni novanta, alla luce di nuove acquisizioni in materia di filogenesi, al punto che i ditterologi che si sono occupati, nell'ultimo ventennio, di queste famiglie (Freidberg, Mathis, Munari, D. McAlpine) hanno frequentemente fatto riferimento all'insieme delle due famiglie usando termini informali come complex o assemblage.
L'unione è stata formalizzata nel 2007 da David Kendray McAlpine. L'entomologo australiano suddivide la famiglia sensu lato nelle sei sottofamiglie che, nell'insieme, componevano i Canacidae sensu stricto e i tradizionali Tethinidae:
Per maggiore chiarezza, in questa sede, si usano le denominazioni Canacidae e Canacidae s.l. (sensu lato) per fare riferimento all'intero taxon secondo la classificazione più recente (sensu D. McAlpine 2007), la denominazione Canacidae s.s. (sensu stricto) per far riferimento all'accezione storica della famiglia con esclusione degli Zaleinae, la denominazione Tethinidae s.s. per far riferimento all'accezione storica della famiglia Tethinidae e, infine, la denominazione Zaleinae per fare riferimento alla sottofamiglia nell'accezione attuale (sensu Freidberg 1995).
Gli adulti hanno corpo di piccole dimensioni, lungo 1,6-5 mm, con livrea di colore variabile dal nero al bruno, al grigio al giallastro e con tegumento di aspetto pruinoso per la copertura tomentosa.
Il capo è grande, di forma variabile ma generalmente globosa, con fronte ampia, gene più o meno ampie, clipeo prominente. È provvisto di tre ocelli e occhi relativamente piccoli. Le antenne sono brevi e aristate, con primo flagellomero di forma globosa e arista pubescente o glabra, inserita in posizione basale e dorsale.
La chetotassi varia secondo le sottofamiglie e si presenta particolarmente complessa a causa di una controversa interpretazione, che riguarda l'identificazione e la denominazione delle setole posizionate dietro il triangolo ocellare. La regione frontale presenta differenti tipi di setole fronto-orbitali:



Le setole ocellari e le verticali esterne ed interne sono bene sviluppate e robuste. Nella regione postocellare è presente, in genere, un paio di setole paraverticali, più o meno sviluppate e robuste, divergenti negli Zaleinae e convergenti nel resto della famiglia, del tutto assenti nei Calacinae e in alcuni Apetaeninae. Queste setole sono inserite dietro il triangolo ocellare e più lateralmente rispetto ai due ocelli posteriori, perciò sono più o meno ampiamente distanziate fra loro. In diversi generi dei Canacinae, in tutte le specie del genere Dasyrhicnoessa e in alcune specie di Tethina, sono presenti altre due setole, posizionate lungo il margine posteriore del triangolo ocellare, fra i due ocelli, e anteriormente rispetto alle altre due paraverticali. Queste due setole sono molto ravvicinate ma largamente divergenti e di sviluppo vario, da sottili a più o meno robuste. Nel resto della famiglia sono invece assenti o insignificanti. La posizione di queste due setole ricorda le cosiddette setole postocellari presenti in molte altre famiglie di Acalyptratae, ma è opinione condivisa che tali setole siano peli ocellari differenziati, perciò sono denominate setole pseudopostocellari, termine proposto da Wirth (1987). Nell'intera famiglia le setole postocellari vere e proprie sarebbero perciò assenti. In realtà questo aspetto è oggetto di controversia fra alcuni Autori, in quanto le setole paraverticali sono ritenute da D. McAlpine vere postocellari. La chetotassi della regione genuale e peristomale comprende annessi tricoidei di vario aspetto secondo il taxon. Le vibrisse sono assenti o presenti, incurvate e più o meno sviluppate, alle quali possono succedere una o più paia di setole sottovibrissali. In diversi Tethinidae s.s. è presente una serie di brevi setole o peli lungo il margine inferiore delle gene, mentre le gene possono glabre o rivestite da radi peli o brevi setole più o meno robuste. Nei Canacinae sono ben visibili una o più setole per lato, lunghe, robuste e inclinate verso l'alto, disposte lungo una linea obliqua che si estende dal margine inferiore dell'occhio ai lati del clipeo.
L'apparato boccale è di tipo lambente-succhiante, ben sviluppato, di lunghezza variabile secondo la conformazione della parte inferiore del capo, piuttosto breve negli Apetaeninae. I labella sono ampi e carnosi e i palpi mascellari moderatamente sviluppati o piuttosto lunghi negli Zaleinae e nei Canacinae.
Il torace è ampio e leggermente convesso. La chetotassi comprende quattro o più paia di setole dorsocentrali, di cui il primo presuturale, 1-2 paia di scutellari, 1-3 paia di omerali (assenti in alcuni Apetaeninae), 1-2 paia di notopleurali e, in genere, 1-3 paia di sopralari, di cui una presuturale, un paio di infralari presuturali e due paia di postalari. Gli annessi acrosticali sono formati da due o più serie di peli più o meno robusti ed eventualmente un paio di setole prescutellari, non sempre presenti. Sulle pleure sono presenti o assenti setole nell'episterno dorsale e nell'episterno ventrale; il numero e la disposizione di queste setole varia secondo il gruppo. Spesso, nell'episterno dorsale sono presenti una o due setole inserite presso il margine posteriore e dirette posteriormente e un'altra setola inserita presso la parte posteriore del margine dorsale, diretta verso l'alto. Le zampe sono relativamente brevi e sottili, rivestite da una pubescenza diffusa, senza setole di particolare sviluppo.
Le ali sono ben sviluppate nella maggior parte della famiglia, generalmente ialine o leggermente offuscate di bruno o giallastro, raramente con pigmentazioni zonali, con alula e lobo anale in genere pronunciati. Alcune specie del genere Apetaenus sono meiottere, ma non si conoscono forme attere. La nervatura è priva di particolari peculiarità. La costa si estende fino alla confluenza della media e presenta una sola frattura subcostale. La subcosta è completa e distinta da R1, ma il suo tratto distale è strettamente ravvicinato a questa e quasi fuso. La radio si divide in tre rami, con R1 breve e robusto e R2+3 e R4+5 lunghi, terminanti in posizione subapicale e apicale. La media è indivisa, la cubito è divisa in due rami, con CuA1 ben sviluppato e CuA2. Quest'ultima è in genere ben conformata ma breve e confluente su A1, mentre negli Zaleinae è quasi del tutto assente. Vena A1+CuA2 sempre incompleta e di sviluppo variabile ma, in generale, è piuttosto breve oppure è prolungata da una leggera piega. La seconda anale (A2) è ben visibile ma non sclerificata. In generale sono presenti e complete le nervature trasversali radio-mediale e medio-cubitale basale e discale, con cellule basali, discale e cup chiuse, ma nei Pelomyiinae manca la medio-cubitale basale, con conseguente fusione della seconda cellula basale con la discale.
L'addome è più o meno cilindrico, composto da sei uriti apparenti nel maschio, sette nella femmina. Nel maschio, gli uriti terminali sono generalmente simmetrici; il sesto tergite si fonde con l'ottavo sternite per formare un unico sclerite dorsale, mentre gli scleriti ventrali sono indistinguibili. Nella femmina, gli uriti terminali sono telescopicamente retrattili nei Tethinidae s.s.. In alcuni generi dei Canacidae s.s., il tergite e lo sternite del settimo urite sono fusi.
Negli Acalyptratae ricorre, in genere, la presenza di due setole, più o meno robuste, posizionate lungo il margine posteriore del triangolo ocellare. Queste setole sono denominate "postverticali" nella vecchia letteratura, già dall'Ottocento e il termine è usato talvolta anche nella letteratura recente. Steyskal (1976) propose la denominazione "setole postocellari"; la stessa è adottata sia nel Manual of Nearctic Diptera[1] sia nel Manual of Palaearctic Diptera[2] e, di conseguenza, ricorre ampiamente nella letteratura che fa riferimento a queste due opere fondamentali. Altre due setole, presenti solo in alcune famiglie di Acalyptratae, sono ubicate posteriormente e lateralmente rispetto al triangolo ocellare, e sono denominate "occipitali interne" nella vecchia letteratura. Steyskal (1976) usa invece la denominazione "setole paraverticali"; la stessa denominazione è adottata nella nomenclatura fondamentale dei due manuali citati.
La natura e la denominazione delle setole paraverticali sensu Steyskal, nei Canacidae, è oggetto di una fondamentale controversia fra J. McAlpine (1989) e D. McAlpine (2007), che discutono in modo dettagliato e approfondito l'omologia, in relazione ad altre famiglie di Acaliptrati, e la conseguente pertinenza della nomenclatura[3][4]. Nel contempo, le principali descrizioni che si sono succedute nel corso di un trentennio, sui singoli taxa che confluiscono nei Canacidae s.l., riportano un quadro complesso e confuso, ai fini della comparazione, per i seguenti motivi:




J. McAlpine (1989), precisando di essere in disaccordo con altri Autori precedenti, fra cui il suo omonimo australiano, ritiene che le due setole esterne e posteriori, presenti nella maggior parte della famiglia e generalmente convergenti, debbano essere interpretate come setole paraverticali sensu Steyskal (1976), denominate anche occipitali interne nella letteratura più datata. Queste setole, pertanto, non sarebbero omologhe delle setole postverticali (o postocellari sensu Steyskal) ricorrenti negli Acalyptratae. Le due setole divergenti più interne, presenti solo in alcuni generi sia dei Tethinidae s.s. sia dei Canacidae s.s. e assenti o insignificanti nel resto della famiglia, vanno interpretate come peli ocellari differenziati e la denominazione più appropriata è quella di setole pseudopostocellari[3].
D. McAlpine (2007), a sua volta, smentisce il suo omonimo canadese e sostiene che le setole più esterne e posteriori (ovvero le paraverticali secondo la terminologia di Steyskal e J. McAlpine) sono in realtà vere setole postverticali e, quindi, omologhe delle setole postocellari usualmente presenti in molti Acaliptrati. In merito alle altre due setole, concorda invece con l'interpretazione proposta da Wirth (1987) e J. McAlpine (1989) nel Manual of Nearctic Diptera, ovvero che trattasi di peli ocellari specializzati e che possano essere denominate setole pseudopostocellari.
In merito alle differenti descrizioni e terminologie adottate, nella tabella seguente sono riassunte le indicazioni riportate da alcuni ditterologi nelle pubblicazioni più significative, citate in letteratura, dalla fine degli anni ottanta ad oggi.
Gli stadi larvali della maggior parte dei Canacidae sono sconosciuti, ad eccezione di poche specie delle sottofamiglie dei Canacinae e degli Apetaeninae. La larva dei Canacinae, nelle forme conosciute, è apoda e microcefala, di forma cilindrica ma affusolata alle due estremità, lunga 5–6 mm, con metameria evidente, di colore biancastro o giallastro. I segmenti addominali, dal secondo al settimo, sono provvisti ciascuno di un'area ambulacrale ventrale prominente, che si estende trasversalmente nella parte anteriore del segmento. Il sistema tracheale, nelle larve di 2ª e 3ª età, è anfipneustico: gli stigmi toracici sono portati da un processo filamentoso retrattile, presente ai lati del protorace, lungo circa un millimetro; quelli addominali sono invece portati all'apice di un tubo respiratorio che emerge dal nono urite, all'estremità caudale.
Il pupario è di colore bruno e ha una forma cilindrica e affusolata, del tutto simile, nell'aspetto, alla larva matura. Sul secondo segmento (primo segmento toracico) sono evidenti i due cornetti respiratori, di maggiore lunghezza rispetto ai processi respiratori della larva.
I Canacidae s.s. sono insetti associati prevalentemente agli ambienti costieri marini: le larve della maggior parte delle specie conosciute sono semiacquatiche e vivono nel piano mesolitorale delle coste, solo poche specie vivono in ambienti d'acqua dolce, rappresentati dalle acque fresche di torrenti montani.
I Tethinidae s.s. sono anch'essi associati ad ambienti costieri, per quanto siano sconosciute le larve della maggior parte delle specie. Gli adulti si rinvengono in genere in ambienti costieri sulla sabbia nuda, su cumuli di alghe e sulla vegetazione delle spiagge o delle retrodune, oppure nelle saline, in stagni e paludi litoranee, nelle mangrovie, sulle rive di laghi salati. Questo comportamento fa presumere che anche le larve dei Tethinidae siano per la maggior parte organismi alofili. Alcune specie dei Tethinidae si rinvengono anche in altri ambienti, come foreste, praterie, deserti, oppure in aree antropizzate, spesso in zone interessate da scarichi industriali o civili.
Il regime dietetico non è conosciuto, ma si presume che sia gli adulti sia le larve si nutrano di detriti organici e alghe[12][13]. Alcune specie dell'areale subantartico sono note per essere associate alle colonie dei pinguini o di altri uccelli marini, in quanto gli adulti sono coprofagi e si nutrono di guano[14][15].
Storicamente, Canacidae s.s. e Tethinidae s.s. sono state trattate come famiglie distinte per lungo tempo. Entrambe furono definite come famiglie da Hendel (1916), rispettivamente con i nomi Canaceidae e Tethinidae[16], Enderlein (1935) usò per la prima volta il nome Canacidae[17]. Per decenni i Canacidae s.s. furono per lungo tempo considerati sistematicamente affini agli Ephydridae, ma tra la fine degli anni cinquanta e gli anni ottanta, fra i ditterologi che si sono occupati di filogenesi degli Schizophora, ha prevalso l'orientamento di correlare i Canacidae alla famiglia dei Tethinidae[18].
La posizione sistematica dei Canacidae s.s. e dei Tethinidae s.s. nei taxa di rango superiore cambia secondo lo schema tassonomico adottato dai diversi Autori. La tradizionale posizione fra gli Acalyptratae era supportata dagli Autori che ritengono monofiletica questa sezione (Hennig, J.F. McAlpine), mentre in altri schemi viene trattata la posizione in una superfamiglia nell'ambito degli Schizophora (Colless & D.K. McAlpine, Griffiths). Colless & D.K. McAlpine (1970) includevano sia i Canacidae sia i Tethinidae nella superfamiglia dei Drosophiloidea[19]. Griffiths (1972), sostenitore della natura parafiletica degli Acalyptratae, inseriva i Tethinidae nei Tephritoinea, una delle otto "prefamiglie" in cui suddivideva la superfamiglia Muscoidea, ma ha trattato i Canacidae come incertae sedis[20]. Hennig (1973) ha incluso sia i Tethinidae sia i Canacidae fra gli Acalyptratae incertae sedis[21]. J.F. McAlpine (1989), che ha dettagliatamente articolato le relazioni filogenetiche fra le famiglie dei Cyclorrhapha, inserisce Tethinidae e Canacidae fra gli Acalyptratae, nella superfamiglia Carnoidea, sulla base del seguente cladogramma[22]:
CarnoideaCryptochetidae+Chloropidae+Milichiidae
Tethinidae
Canacidae (incluso Zalea)
La relazione filogenetica tra Tethinidae e Canacidae s.s è sostenuta da McAlpine per le seguenti sinapomorfie:
Come autapomorfie proprie dei Canacidae, McAlpine segnala la direzione lateroclinata (ovvero inclinata all'esterno) delle setole fronto-orbitali inferiori, la faccia prominente, le gene gene provviste di setole genuali incurvate verso l'alto, il clipeo largo, la presenza del ponte ipandriale provvisto di spine e l'edeago breve nel maschio e la conformazione dei cerci nella femmina, sclerificati, ricurvi e provvisti di 1-2 spine apicali.
La stretta affinità filogenetica faceva ipotizzare, già dagli anni ottanta, che Canacidae e Tethinidae potessero essere riuniti in un'unica famiglia. Questa ipotesi era prospettata sia da David Kendray McAlpine sia da James Francis McAlpine, tuttavia entrambi trattarono come distinte le due famiglie a causa di insufficienti elementi che potessero avallare l'unione[18]. Freidberg (1995) ha definito un nuovo genere, Suffomyia, inserendolo nella sottofamiglia Zaleinae[8]. In seguito all'aggiunta di Suffomyia, Freidberg ha ridefinito le relazioni filogenetiche fra le due famiglie arrivando alla conclusione che la sottofamiglia degli Zaleinae si colloca in una posizione intermedia fra i Tethinidae s.s. e i Canacidae s.s. e che non può trovare collocazione né fra i Canacidae né fra i Tethinidae. L'entomologo israeliano formulò perciò la proposta di unione dei tre taxa in un'unica famiglia che avrebbe dovuto prendere il nome di Canacidae, secondo le convenzioni del Codice Internazionale di Nomenclatura Zoologica[8][23][24]. Pochi anni più tardi, Munari (1998) tratta i Tethinidae, nel Manual of Palaearctic Diptera, includendovi formalmente la sottofamiglia Zaleinae[25].
L'unione delle due famiglie è formalmente definita da D.K. McAlpine (2007). L'entomologo australiano ipotizza, per la nuova famiglia, tre autapomorfie[26]:
D. McAlpine, inoltre, adotta come sottogruppi le sottofamiglie definite in precedenza per suddividere le famiglie dei Canacidae e dei Tethinidae e vi posiziona i vari generi, individuando per ciascuna di esse le autapomorfie specifiche e le apomorfie condivise con altre sottofamiglie. La sola collocazione incerta riguarda il genere Plesiotethina, che McAlpine inserisce con riserva nella sottofamiglia Tethininae. Infine, l'Autore elabora le chiavi di determinazione tassonomica delle sottofamiglie e, approfondendo la trattazione degli Zaleinae, le chiavi di determinazione di generi e specie di quest'ultima sottofamiglia.
A distanza di pochi anni, la revisione sistematica di D.K. McAlpine trova ancora pochi riscontri, almeno nella tassonomia adottata nei cataloghi on line e nei principali siti che trattano dell'ordine dei Ditteri. Fra questi, il BioSystematic Database of World Diptera ha aggiornato la lista delle famiglie, riportando il nome Tethinidae come sinonimo di Canacidae, tuttavia non ha ancora aggiornato il database, attribuendo separatamente le varie specie alle vecchie famiglie sensu stricto.
Nel complesso, i Canacidae sensu lato comprendono oltre 300 specie ripartite fra sei sottofamiglie e 28 generi[27][28][29][30]:
La famiglia ha un'ampia distribuzione ed è rappresentata in tutte le ecozone, ma con distribuzioni differenti secondo la sottofamiglia[27]:
Un aspetto particolare relativo alla distribuzione mondiale dei Canacidae è la segnalazione della presenza di almeno 13 specie nelle Isole Galapagos, la maggior parte presumibilmente endemiche. Queste specie sono Canacea snodgrasii, Canaceoides angulatus, Nocticanace ashlocki, Nocticanace cancer, Nocticanace curioi, Nocticanace galapagenis, Nocticanace darwini, Nocticanace scapania, Nocticanace spinicosta, Nocticanace usingeri, Paracanace cavagnaroi, Paracanace maritima[29] e Tethina insulans[32].
In Europa sono segnalate circa 40 specie appartenenti ai generi Tethina, Pelomyiella, Pelomyia, Canace e Xanthocanace, ma tre quarti di queste fanno capo al genere Tethina[33][34].
In Italia sono segnalate poco meno di venti specie, quasi tutte appartenenti al genere Tethina; le restanti appartengono ai generi Canace e Pelomyiella[35][36]. La presenza di alcune specie nel territorio italiano e l'effettiva distribuzione di diverse Tethina sono tuttavia incerte.
Canacidae Jones, 1906, è una famiglia di insetti dell'ordine dei Ditteri (Brachycera: Cyclorrhapha: Acalyptratae), simili nell'aspetto ai moscerini della frutta.
La maggior parte di questi insetti è associata ad ambienti costieri marini ed oceanici, perciò sono noti, in inglese, con nomi che fanno riferimento al loro tipico habitat, come beach flies, surf flies e surge flies ("mosche della spiaggia", "dell'onda", "della marea").
Canacidae zijn een familie uit de orde van de tweevleugeligen (Diptera), onderorde vliegen (Brachycera). Wereldwijd omvat deze familie zo'n 28 genera en 323 soorten. In Nederland is één soort inheems, de Xanthocanace ranula.
Canacidae zijn een familie uit de orde van de tweevleugeligen (Diptera), onderorde vliegen (Brachycera). Wereldwijd omvat deze familie zo'n 28 genera en 323 soorten. In Nederland is één soort inheems, de Xanthocanace ranula.
Canacidae, eller canacider som de kalles på norsk, er en liten familie av fluer som lever i tare og andre oppskylte alger på havstrender. De ligner Tethinidae, som de er nært beslektet med, men skiller seg fra disse ved å være kraftigere bygd og mer hårete. De fleste artene lever i varme klima, to arter finnes i Nord-Europa men det er usikkert om noen av dem når så langt nord som til Norge.
Små til middelsstore (1,5 – 5 mm) fluer, ofte dekket av lys bestøvning. De ligner på Tethinidae men er kraftigere og mer hårete. Brystet (thorax) har både korte hår og lange børster. Bakkroppen er sylindrisk.
Hodet har tre-fire par kraftige, utoverrettede børster ved innerkanten av hvert fasettøye. Kinnene er høye som hos Tethinidae, vanligvis med tre børster som er mye kraftigere enn den øvrige behåringen. De har vanligvis to rekker av fine børster i pannen, og et eller to par kraftige børster nær punktøynene.
Vingene er klare uten fargeflekker. Beina er slanke, tett hårete men uten kraftige børster.
Larvene er pølseformede, tilspisset i begge ender. Bakenden er trukket ut til et kort ånderør. Hodeenden er moderat tilspisset. På undersiden av hvert kroppssegment er det noen tverr-rader av korte kroker som hjelper dem feste seg til underlaget.
De fleste artenes larver utvikler seg trolig i råtnende tang, tare og andre alger som ligger oppskyllet ved havet. Noen ganske få arter lever ved ferskvann, deres biologi er ukjent. De fleste av artene i denne familien lever i varme klima, for eksempel ved Middelhavet.
Canacidae, eller canacider som de kalles på norsk, er en liten familie av fluer som lever i tare og andre oppskylte alger på havstrender. De ligner Tethinidae, som de er nært beslektet med, men skiller seg fra disse ved å være kraftigere bygd og mer hårete. De fleste artene lever i varme klima, to arter finnes i Nord-Europa men det er usikkert om noen av dem når så langt nord som til Norge.
Canacidae Jones, 1906
СинонимыCanacidae (лат.) — семейство насекомых из отряда двукрылых. В семействе описано около 300 видов и 27 родов (6 подсемейств)[1]. Мухи очень мелких размеров; в длину достигают всего 1,6—5 мм. Личинки развиваются в вянущих водорослях, которых выбросило на берег. Встречаются в регионах с тёплым климатом, главным образом, в Средиземноморье.
Canacidae (лат.) — семейство насекомых из отряда двукрылых. В семействе описано около 300 видов и 27 родов (6 подсемейств). Мухи очень мелких размеров; в длину достигают всего 1,6—5 мм. Личинки развиваются в вянущих водорослях, которых выбросило на берег. Встречаются в регионах с тёплым климатом, главным образом, в Средиземноморье.

Canaceidae Hendel, 1916
英名 beach fliesニセミギワバエ科(Canacidae)は、ハエ目(双翅目)に属する科の一つ。ニセミギワバエ科に属する種は世界中に分布しており、ニセミギワバエ Procanace grisescens など307種が記録されている[1]。
和名はイソバエ科ともいう。
ニセミギワバエ科の多くの種は海岸近くなどで見られるが、内陸の塩生の土地やアルカリ性の環境に生息する種もある[1]。またハワイでは草原や淡水域にみられる種もある[1]。
ニセミギワバエ科の種の体サイズは1.6-4.0mmで、小型種または中型種である[1]。体は薄い黄色から褐色の細かい毛でおおわれていることが多い[1]。
成虫の外見はミギワバエ科の各種と類似するが、翅の臀室や第一経脈の有無(ミギワバエにはどちらもない、ニセミギワバエにはどちらもある)、前縁脈の切れ込み位置の違いなど、翅脈によって識別可能である[2]。
ニセミギワバエ科には6つの亜科が含まれるが、そのうち Tethininae 亜科はハマベホソバエ科(イソベバエ科、Tethinidae)として独立の科として扱われていた。しかし2000年代以降の研究によって、ハマベホソバエ科は側系統群であることが示唆され、ハマベホソバエ科を広義のニセミギワバエ科(Canacidae s.l.)に含めるとする主張が示されている[1]。
ニセミギワバエ科に属する分類群として6亜科27属307種が記録されているが[1]、ハマベホソバエ科を独立の科として扱う場合、約16属180種がハマベホソバエ科に属する。