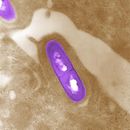
Listeria monocytogenes é unha especie de bacterias grampositivas que causa unha infección chamada listeriose. É un organismo anaerobio facultativo, que pode sobrevivir en presenza ou en ausencia de oxíxeno. Pode crecer e reproducirse dentro das células hóspede e é un dos patóxenos máis virulentos que poden estar presentes nos alimentos, ata o punto de que entre o 20 e o 30 % das infeccións clínicas por esta bacteria acaban sendo mortais.[1] A listeriose é a principal causa de morte por infeccións alimentarias, superando en taxas de mortalidade a Salmonella e Clostridium botulinum.
Presenta motilidade por medio de flaxelos a 30 °C ou menos, pero xeralmente non a 37 °C,[2] L. monocytogenes pode moverse entre as células eucariotas sen usar os flaxelos pola polimerización explosiva de filamentos da proteína actina, o que se coñece como "colas de cometa" ou "foguetes de actina".
Algúns estudos suxiren que ata o 10% do tracto gastrointestinal humano pode estar colonizado por L. monocytogenes.[1] Con todo, as enfermidades clínicas debidas a L. monocytogenes son máis frecuentes en veterinaria que en medicina humana, especialmente as meningoencefalites en ruminantes.
Debido á súa frecuente patoxenicidade, que causa meninxite en neonatos (adquirida transvaxinalmente), as nais preñadas deben ter coidado ao comer queixos que poidan estar contaminados coa bacteria.[3] É a terceira causa máis común de meninxite en neonatos.
L. monocytogenes é unha bacteria grampositiva, non formadora de esporas, móbil, anaerobia facultativa, e de forma bacilar (0,4 a 0,5 microns x 0,5 a 1,2). É catalase positiva e oxidase negativa, e expresa unha beta hemolisina, que causa a destrución dos glóbulos vermellos do sangue. Mostra unha característica motilidade dando voltas ollada con microscopio óptico.[4] Aínda que L. monocytogenes é activamente móbil utilizando os seus flaxelos peritricos a temperatura entre 20−25 °C, pero o organismo non utiliza flaxelos á temperatura corporal de 37 °C.[5]
O xénero Listeria pertence á clase Bacilli, e á orde Bacillales, na que tamén están incluídos os xéneros Bacillus e Staphylococcus. das especies clasificadas no xénero Listeria hai dúas, L. ivanovii e L. monocytogenes, que son patóxenas de animais, como os ratos, pero só L. monocytogenes está demostrado que produce enfermidades en humanos.[6] Hai 13 serotipos de L. monocytogenes que poden causar enfermidades, pero máis do 90 % dos illados humanos pertencen só a tres deles, que son: 1/2a, 1/2b, e 4b. As cepas do serotipo 4b de L. monocytogenes son responsables dos principais casos de enfermidades de orixe alimentaria humanas en Europa e Norteamérica e de entre o 33 e o 50 % dos casos esporádicos no mundo desde a década de 1980.[7]
O nome do xénero Listeria púxose en honor do médico británico Joseph Lister. A especie L. monocytogenes foi descrita por primeira vez por E.G.D. Murray en 1926 baseándose en seis casos de mortes repentinas de crías de coellos.[8] Murray denominou ao organismo Bacterium monocytogenes pero despois Harvey Pirie cambioulle o nome polo de Listeria en 1940.[9] O epíteto monocytogenes débese a que extractos da súa membrana poden estimular (xerar) a produción de monocitos no coello, aínda que non no ser humano.[10] Aínda que as descricións clínicas da infección por L. monocytogenes en animais e humanos se publicaron na década de 1920, ata 1952 en Alemaña Oriental non foi recoñecida como unha causa significativa de sepse e meninxite neonatal.[11] A listeriose en adultos sería despois asociada con pacientes con sistemas inmunitarios comprometidos, como os que toman fármacos inmunosupresores e corticosteroides para tratar tumores malignos ou transplantes de órganos, e tamén os que teñen infección por VIH/SIDA.[12]
Ata 1981 L. monocytogenes non foi identificada como causa de enfermidades de orixe alimentaria. Naquel ano un estalido de listeriose na cidade de Halifax, Nova Escocia na que se produciron 41 casos e 18 mortes, principalmente en mulleres preñadas e neonatos, foi ligada epidemioloxicamente co consumo de ensalada de col que fora fertilizada con esterco de ovella contaminado con L. monocytogenes.[13] Desde entón, L. monocytogenes é recoñecida como un importante perigo na industria da alimentación.[14]
A infección invasiva por L. monocytogenes causa a enfermidade chamada listeriose. Cando a infección non é invasiva, a enfermidade consecuencia desta infección denomínase gastroenterite febril. Entre as manifestacións da listeriose están a septicemia,[15] meninxite (ou meningoencefalite),[15] encefalite,[16] úlcera corneal,[17] pneumonía,[18] e infeccións intrauterinas ou de cérvix en mulleres embarazadas, o que pode orixinar aborto espontáneo (no segundo ou terceiro trimestre) ou morte fetal. Os neonatos que sobreviven poden sufrir unha granulomatose (granulomas pióxenos distribuídos por todo o corpo) e atraso físico. Estes trastornos son xeralmente precedidos por síntomas parecidos aos dunha gripe, como febre persistente. Os síntomas gastrointestinais, como náuseas, vómitos e diarreas, poden preceder a formas máis serias de listeriorse ou poden ser os únicos síntomas.
Un estudo inicial suxeriu que L. monocytogenes era a única bacteria grampositiva que posuía lipopolisacárido,[19] que lle servía como endotoxina, pero posteriormente viuse que non era unha verdadeira endotoxina. As paredes celulares da bacteria sempre conteñen ácidos lipoteicoicos, nos cales un resto glicolípido, como o galactosil-glicosil-diglicérido, está ligado covalentemente ao fosfomonoéster terminal do ácido teicoico. Esta rexión lipidica ancora a cadea do polímero á membrana plasmática. Estes ácidos lipoteicoicos lembran aos lipopolisacáridos das bacterias gramnegativas en estrutura e función, e son os únicos polímeros anfipáticos na superficie celular.[20][21]
L. monocytogenes ten residuos de D-galactosa na súa superficie que poden unirse a receptores de D-galactosa na superficie da célula hóspede. Estas células hóspede adoitan ser as células M e as das placas de Peyer da mucosa intestinal. Unha vez unidas a estas células, L. monocytogenes pode atravesar a mucosa intestinal e entrar no corpo.
A dose infectiva de L. monocytogenes varía segundo a cepa e a susceptibilidade da vítima. Nos casos contraídos a partir de leite sen pasteurizar ou supostamente pasteurizado, pódese asumir que en persoas susceptibles, abonda con menos de 1.000 organismos totais para causar enfermidade. L. monocytogenes pode invadir o epitelio gastrointestinal. Unha vez que a bacteria entra nos monocitos, macrófagos, ou leucocitos polimorfonucleares do hóspede, convértese nunha septicemia no sangue e pode proliferar. A súa presenza intracelular en células fagocíticas tamén lle permite acceder ao cerebro e probablemente facer unha migración transplacentaria ata o feto nas mulleres embarazadas. A patoxénese de L. monocytogenes céntrase na súa capacidade de sobevivir e multiplicarse nas células fagocíticas. Parece que esta bacteria inicialmente evolucionou para poder invadir as mucosas intestinais como unha infección intracelular, e desenvolveu un mecanismo químico para facelo no que intervén a proteína bacteriana internalina, que se une á proteína cadherina da membrana plasmática da célula intestinal. Estas moléculas de adhesión tamén se encontran noutras dúas robustas barreiras do coropo humano, como son a barreira hematoencefálica e a barreira placentaria, e isto pode explicar a aparente afinidade de L. monocytogenes por causar meninxites e afectar aos fetos.
L. monocytogenes pode actuar como saprófita ou como patóxena, dependendo das condicións do seu ambiente. Cando a bacteria está presente nun organismo hóspede, o fenómeno da percepción do quórum ou quorum sensing causa a regulación á alza de varios xenes de virulencia. Dependendo da localización da bacteria no organismo hóspede, diferentes activadores regulan á alza os xenes da virulencia. O factor SigB é un factor sigma alternativo que regula os xenes Vir nos intestinos, mentres que o PrfA regula á alza a expresión de xenes cando a bacteria se encontra no sangue.[22][23][24][25] Pouco se sabe sobre como esta bacteria cambia de actuar como saprófita a como patóxena; porén, pénsase que son necesarios varios ARNs non codificantes de L. monocytogenes para inducir este cambio.
L. monocytogenes ten tres liñaxes, con historias evolutivas e potenciais patoxénicos diferentes.[26] As cepas da liñaxe I son a maioría dos illados clínicos humanos e todos os clons epidémicos humanos, pero están infrarrepresentados nos illados clínicos animais.[26] As cepas da liñaxe II están sobrerrepresentados nos casos animais e todo o contrario nos casos clínicos humanos, e son máis prevalentes en mostras tomadas do ambiente e de alimentos.[27] Os illados da liñaxe III son moi raros, pero significativamente máis comúns nos animais que nos illados humanos.[26]
Proba de Anton: é unha proba usada na identificación de L. monocytogenes; a instilación dun cultivo en saco conxuntivo de ollo de coello ou coello de Indias causa unha grave queratoconxuntivite en 24 horas.[29][30]
Cultivo: Listeria crece en medios como o ágar de Mueller-Hinton. A identificación vese potenciada se os cultivos primarios se fan en ágar que conteña sangue de ovella, porque pode observarse a pequena zona característica de hemólise arredor e baixo as colonias. O illamento pode mellorarse se o tecido de procedencia se mantén a 4 °C durante algúns días antes de facer a inoculación no medio de cultivo bacteriolóxico. O organismo é un anaerobio facultativo e é catalase positivo e motil. Produce ácido pero non gas con diversos carbohidratos.[31] A motilidade á temperatura dunha habitación e a produción de hemolisina son datos fundamentais para diferenciala das bacterias corineformes.
Ágar bile esculina. Desenvólvese ben en bile, polo que se utilizan medios inclinados con ágar bile esculina. Esta proba consiste en determinar a capacidade que ten L. monocytogenes de hidrolizar a esculina a esculetina e a glicosa en presenza de sales biliares. A esculetina xerada reacciona cos ións de ferro que contén o cloruro férrico no medio e xera unha coloración negra, se é positiva.[32]
Reacción de Voges-Proskauer. L. monocytogenes presenta un metabolismo fermentativo que xera ácido a partir da glicosa e produce acetona, o que dá lugar a unha reacción de Voges-Proskauer positiva e non fermenta a xilosa. A fermentación destes azucres ponse en evidencia pola técnica de Voges-Proskauer, a cal consiste en identificar se L. monocytogenes xera ácidos e diacetil, xa que o medio contén azucres fermentables. Para ver a reacción utilízanse reveladores como alfa-naftol e KOH. Se a suspensión se torna de cor rosa a proba é positiva.[32]
Proba de CAMP. A proba de CAMP consiste en sementar en ágar sangue unha estría en forma horizontal de L. monocytogenes e unha perpendicular a esta de Staphylococcus aureus sen que se unan entre si. O resultado agardado será que L. monocytogenes xere o factor CAMP, o cal produce un sinerxismo coa beta lisina producida por S. aureus sobre os eritrocitos, xerando a lise destes.[33]
Os métodos de análise de alimentos son complexos e levan moito tempo (varios días, ata sete), pero o anuncio da aparición de novas probas que usan sondas de ADN específicas non marcadas radioactivamente debería permitir unha confirmación máis rápida e simple dos illados sospeitosos. A tecnoloxía do ADN recombinante pode tamén permitir unha análise en dous ou tres días no futuro.
Cando se produce meninxite por L. monocytogenes, a taxa de mortalidade pode ser do 70%, na septicemia do 50%, e nas infeccións neonatais maior do 80%. Nas infeccións durante o embarazo a nai normalmente sobrevive. Hai informes de que funciona o tratamento con penicilina ou ampicilina e trimetoprim-sulfametoxazol.
Propúxose o uso do bacteriófago chamado fago de Listeria P100 como aditivo alimentario para controlar a Listeria monocytogenes.[34] Varias compañías (EBI Food Safety, Intralytix) desenvolveron tratamentos para os alimentos con bacteriófagos, que se aplicarían sobre froitas e carnes como o xamón e o pavo.
Listeria é resistente ás penicilinas naturais (penicilina G, penicilina V), penicilinas resistentes a β-lactamase (meticilina, nafcilina, isoxazoilpenicilinas —oxacilina, cloxacilina, dicloxacilina—) e a todas as cefalosporinas. Pero é susceptible ás penicilinas de espectro ampliado.[35]
Como L. monocytogenes é unha bacteia intracelular, algúns estudos utilizárona como vector para transportar xenes ao interior de células in vitro. Actualmente a eficiencia desta transfección non é moi grande. Un exemplo do uso con éxito de L. monocytogenes en tecnoloxías de transfección in vitro é na terapia xénica para casos de fibrose quística.[36]
Estase a desenvolver unha vacina contra o cancro viva atenuada de L. monocytogenes ADXS11-001 como posible tratamento do carcinoma cervical.[37]
L. monocytogenes atopouse en polo menos 37 especies de mamíferos, tanto domésticos coma salvaxes, e en polo menos 17 especies de aves e posiblemente nalgunhas especies de peixes e mariscos. L. monocytogenes pode illarse do solo, ensilados, e outras fontes ambientais. L. monocytogenes é bastante resistente e aguanta os efectos deletéreos da conxelación, desecación, e da calor notablemente ben para unha bacteria que non forma esporas. A maioría das cepas de L. monocytogenes son patóxenas nalgún grao.
L. monocytogenes foi asociada con alimentos como o leite (xeralmente sen pasteurizar),[38] queixos (especialmente as variedades brandas curadas), xelados, vexetais crus, salchichas de carne crúa fermentada, carne de aves crúa ou cociñada, carnes crúas de todos os tipos, e peixe cru e afumado. A súa capacidade de crecer a temperaturas frías de ata 0 °C permite a súa multiplicación en alimentos refrixerados. Á temperatura de refrixeración de 4 °C, a cantidade de ferro férrico pode afectar ao crecemento de L. monocytogenes.[39]
O sitio principal de infección é o epitelio intestinal, no cal a bacteria invade as células non fagocíticas por medio dun mecanismo de cremalleira. A captación é estimulada pola unión das internalinas de listeria (Inl) ás E-cadherinas (factor de adhesión) da célula hóspede, ou ao factor Met (c-Met, factor de crecemento do hepatocito). Esta unión activa certas GTPases Rho, que seguidamente se unen e estabilizan á proteína da síndrome de Wiskott Aldrich (WAsp). A WAsp pode despois unirse ao complexo Arp2/3 e servir como un punto de nucleación da actina. Despois, a polimerización da actina crea unha "copa fagocítica", é dicir, unha estrutura baseada na actina que normalmente se forma nos fagocitos arredor de materiais alleos antes da endocitose. O efecto neto da unión da internalina é aproveitar esta formación do hóspede para a entrada da bacteria. L. monocytogenes pode tamén invadir as células fagocíticas (por exemplo, os macrófagos), pero só precisa das internalinas para a invasión das células non fagocíticas.
Despois da súa entrada na célula, a bacteria pode escapar do vacúolo/fagosoma antes de que se una cun lisosoma. Tres dos principais factores de virulencia que lle permiten á bacteria escapar do fagosoma son a listeriolisina O (LLO, codificada polo xene hly), a fosfolipase A (codificada polo xene plcA) e a fosfolipase B (codificada no plcB).[40][41] A secreción de LLO e o produto de PlcA distorsionan a membrana vacuolar e permiten á bacteria escapar ao citoplasma, onde pode proliferar.
Unha vez no citoplasma, L. monocytogenes aprovéitase da actina do seu hóspede por segunda vez. As proteínas ActA asociadas co polo vello da célula bacteriana (ao ser un bacilo, L. monocytogenes forma un septo durante a división no centro da célula, onde se forma o polo novo, e así dise que ten un polo novo e un polo vello) poden unirse ao complexo Arp2/3, inducindo así a nucleación da actina nunha área específica da superficie da célula bacteriana. A polimerización da actina despois propulsa á bacteria unidireccionalmente na membrana plasmática do hóspede. A protrusión que se forma pode despois ser captada por unha célula veciña que a introduce no seu interior, formando un vacúolo de dobre membrana do cal a bacteria debe escapar usando a LLO e a PlcB. Este modo de diseminación directo de célula a célula implica un mecanismo celular coñecido como paracitofaxia.[42]
Listeria monocytogenes é unha especie de bacterias grampositivas que causa unha infección chamada listeriose. É un organismo anaerobio facultativo, que pode sobrevivir en presenza ou en ausencia de oxíxeno. Pode crecer e reproducirse dentro das células hóspede e é un dos patóxenos máis virulentos que poden estar presentes nos alimentos, ata o punto de que entre o 20 e o 30 % das infeccións clínicas por esta bacteria acaban sendo mortais. A listeriose é a principal causa de morte por infeccións alimentarias, superando en taxas de mortalidade a Salmonella e Clostridium botulinum.
Presenta motilidade por medio de flaxelos a 30 °C ou menos, pero xeralmente non a 37 °C, L. monocytogenes pode moverse entre as células eucariotas sen usar os flaxelos pola polimerización explosiva de filamentos da proteína actina, o que se coñece como "colas de cometa" ou "foguetes de actina".
Algúns estudos suxiren que ata o 10% do tracto gastrointestinal humano pode estar colonizado por L. monocytogenes. Con todo, as enfermidades clínicas debidas a L. monocytogenes son máis frecuentes en veterinaria que en medicina humana, especialmente as meningoencefalites en ruminantes.
Debido á súa frecuente patoxenicidade, que causa meninxite en neonatos (adquirida transvaxinalmente), as nais preñadas deben ter coidado ao comer queixos que poidan estar contaminados coa bacteria. É a terceira causa máis común de meninxite en neonatos.
_409-14,_Figure_1.png)
