Els mimivirus són un gènere viral (no tothom està d'acord en aquest punt) que conté una sola espècie identificada: Acanthamoeba polyphaga mimivirus (APMV). Té la càpsida de diàmetre més gros conegut en cap virus, així com el genoma més gran i complex que s'hagi pogut trobar mai en un virus. Tot i que el coneixement sobre els virus és relativament limitat, el descobriment d'aquest causà un cert enrenou degut a les implicacions de la seva complexa natura, havent-hi gent proclamant que hauria de donar lloc a un nou domini de la vida intermedi entre els virus i Cytota.
L'APMV va ser descobert per fortuïtament el 1992 en l'ameba Acanthamoeba polyphaga per raó de la qual va ser anomenada, mentre on efectuava un treball de recerca sobre la Legionellosi. El virus va ser observat en una tinció de gram i es va confondre per error amb un bacteri gram positiu. Conseqüentment va ser anomenat "Bradfordcoccus", ja que la font de l'ameba era Bradford, a Anglaterra. El 2003, investigadors de la Université de la Méditerranée de Marsella, a França, varen publicar un article a Science identificant l'organisme com a virus.[1]
El Comitè Internacional per a la Taxonomia de Virus encara no l'ha col·locat en cap família vírica, però sí que s'ha proposat la família Mimiviridae que per metagenòmica es creu que tindrà més espècies.[2] S'ha col·locat tanmateix en el Grup I de la classificació de Baltimore.
Encara que no és estrictament un mètode de classificació s'ha col·locat en un grup de virus grossos consegut com a virus nucleoplasmàtics d'ADN (NCLDV), que inclou altres quatre famílies: Poxviridae, Iridoviridae, Phycodnaviridae i Asfarviridae. És un grup de virus que comparteixen característiques moleculars i grans genomes. El genoma del mimivirus també posseeix 21 gens que codifica per proteïnes homòlogues que s'han trobat altament conservades de la majoria de NCLDVs. Treballs addicionals han suggerit que va divergir en estadis primerencs de l'establiment del grup dels NCLDV.[1]
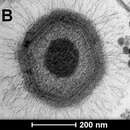
Els mimivirus són els virus més grans coneguts, amb un diàmetre de la càpsida de 400 nm. Tenen filaments de proteïna que projecten 100 nm des de la superfície de la càpsida fent que el total de la seva llargada sigui de 600 nm. En les fonts científiques la mida del virió oscil·la entre els 400 i 800 depenent de com i què es mesuri. La càpsida s'ha vist per microscopi electrònic que és icosaèdrica. No s'ha pogut observar una envolta viral, suggerint que el virus no surt de la cèl·lula per exocitosi.[3]
Els mimivirus comparteixen diverses característiques morfològiques comunes amb tots els membres del grup NCLDV dels virus. Com que tota la resta de virus d'aquest grup contenen una capa lipídica envoltant el nucli central M.Suzan-Monti et al. ha suggerit que també pot ser present en els mimivirus. En el microscopi electrònic apareix com a una regió fosca el nucli central del virió. El genoma del virus es troba en aquesta àrea.
Es poden recuperar diversos ARNm dels virions purificats. S'hi ha trobat el factor de transcripció filo-TFII i una proteïna de càpsida com en altres NCLDV. Tanmateix també s'hi ha trobat tres transcrits d'enzims d'aminoacil tRNA sintetase i quatre molècules desconegudes específiques de mimivirus. Aquests transcrits preempaquetats es poden traduir sense expressió genètica viral i semblen necessaris per al Mimivirus per a la replicació. Altres virus d'ADN com el citomegalovirus humà i l'Herpes simplex virus tipus-1, que també tenen la característica de posseir transcrits d'ARNm preempaquetats (M. Suzan-Monti, 2006).
El genoma del mimivirus és una molècula d'ADN linear i de doble catenària que fa uns 1,2 milions de parells de bases. Això el converteix en el virus amb el genoma més gros conegut per la ciència, doblant el següent virus en llargada conegut, el myovirus Fag G de Bacillus. És fins i tot més gros que una trentena d'organismes cel·lulars coneguts.[4]
A més del genoma tan gros s'estima que els mimivirus codifiquen per a unes 911 proteïnes, quantitat molt més elevada que les quatre amb les que s'ha provat que un virus és viable (Vegeu MS2 and Qβ viruses[5]). L'anàlisi del seu genoma revela la presència de gens no vistos en altres virus, incloent-hi l'aminoacil tRNA sintetasa, i altres gens que es creia que només codificaven organismes cel·lulars. Com altres virus d'ADN, conté diversos gens per al metabolisme dels sucres, dels lípids i els aminoàcids, així com d'altres gens del metabolisme no trobats en altres virus (M. Suzan-Monti, 2006). Aproximadament un 90% del genoma té capacitat codificant mentre que el 10% restant s'ha classificat com a ADN escombraries.
Els estadis de la replicació dels mimivirus no són massa coneguts, però se sap com a mínim que ataca un receptor específic de la superfície de l'ameba la qual cosa l'introdueix cap a l'interior de la cèl·lula. Un cop dins ocorre la fase d'eclipsi en la que el virus no es fa perceptible al microscopi. Després d'unes quatre hores es comencen a contemplar acumulacions en algunes àrees de la cèl·lula. Vuit hores després de la infecció es fan visibles els virions encara dintre de la cèl·lula. El citoplasma de la cèl·lula es continua emplenant amb nous virions i 24 hores després de la infecció inicial la cèl·lula esclata i allibera els nous virions (M. Suzan-Monti, 2006).
Se sap poc d'aquest cicle replicatiu. Amb la microfotografia s'ha pogut establir, però, que els mimivirus s'encapsiden en el nucli, l'adquisició d'una membrana lipídica interna fent servir la del nucli, i partícules semblants trobades en diversos virus, incloent-hi tot els membres de NCLDV. Aquestes partícules són conegudes en altres virus com a fàbriques virals i permeten una encapsidació eficient modificant amples àrees de la cèl·lula hoste.
Els Mimivirus posseeixen diverses característiques que els situen en el llindar del que separa vida i matèria inerta. Són tan grans com diverses espècies bacterianes anomenades Rickettsia conorii i Tropheryma whipplei, posseeixen un genoma comparable en mida a diversos bacteris, incloent-hi aquests dos mateixos, i codifiquen per a productes que abans es creia que no codificaven els virus. A més, els virus posseeixen gens que codifiquen per a nucleòtids i la síntesi d'aminoàcids que fins i tot manquen en alguns bacteris intracel·lulars estrictes. Això significa que, a diferència d'aquests bacteris, els Mimivirus no tenen dependència de l'expressió d'aquestes molècules en la cèl·lula que parasiten. Tanmateix els manquen els gens de les proteïnes ribosòmiques, fent-los dependents de la maquinària de traducció i el metabolisme energètic. Aquests factors combinats han portat els científics al debat de si els Mimivirus podrien ser considerats una forma diferent de vida, comparable en escala als dominis Eukarya, Archaea i Eubacteria.
Com que el seu llinatge és molt antic i que podria haver emergit abans que els organismes cel·lulars, el coneixement sobre els Mimivirus ha portat informació sobre l'origen de la vida. Alguns gens únics dels Mimivirus, incloent-hi els que codifiquen per a la càpsida, han estat conservats en una varietat de virus que infecten organismes de tots els dominis - Eukarya, Archaea i Bacteria. Això s'ha fet servir per suggerir que estan relacionats a un tipus de virus d'ADN que varen emergir abans de la vida cel·lular i que varen jugar un paper important en el desenvolupament de la vida a la terra.[6] Una hipòtesi alternativa és que hi va haver tres tipus diferents de virus d'ADN que es varen veure implicats en la generació dels tres dominis de la vida.[7]
Els mimivirus són un gènere viral (no tothom està d'acord en aquest punt) que conté una sola espècie identificada: Acanthamoeba polyphaga mimivirus (APMV). Té la càpsida de diàmetre més gros conegut en cap virus, així com el genoma més gran i complex que s'hagi pogut trobar mai en un virus. Tot i que el coneixement sobre els virus és relativament limitat, el descobriment d'aquest causà un cert enrenou degut a les implicacions de la seva complexa natura, havent-hi gent proclamant que hauria de donar lloc a un nou domini de la vida intermedi entre els virus i Cytota.
Mimivirus je rod virů s dvouvláknovou DNA, výjimečný svými rozměry (průměr dvacetistěnné kapsidy 400 nm, celkově až 800nm) i výjimečně velkým genomem (1 181 404 párů bází).[1][2] Je znám jediný druh – Acanthamoeba polyphaga mimivirus (APMV). Virus byl nalezen v měňavkovci Acanthamoeba polyphaga a pro svou velikost byl zprvu považován za bakterii.
Mimivirus patří do skupiny tzv. jaderně-cytoplazmatických virů s velkou DNA označované NCLDV (z anglického nucleocytoplasmic large DNA viruses). Ta je někdy považována za novou, čtvrtou doménu živých organismů. Právě velký genom virů rodu mimivirus jako typických zástupců skupiny umožnil provést molekulárně biologické srovnání s ostatními doménami, poskytující indicie pro fylogenetické vyčlenění této domény.[3][4] Existence nové domény je zatím přijímána skepticky.[5][6]
Do příbuznosti mimiviru je řazen i rekordman ve velikosti virů z r. 2011 – Megavirus chilensis. Jeho genom je ještě o 6,5 % větší než genom mimiviru – obsahuje 1 259 197 párů bází.[7][8] V roce 2013 pak rekord převzaly pandoraviry, pravděpodobně také z příbuznosti mimiviru – konkrétně Pandoravirus salinus obsahuje 2,47 milionů párů bází a s velikostí 1 mikrometr je dokonce viditelný optickým mikroskopem.[9][10] V roce 2014 pozici rekordmana převzal ještě o polovinu větší pithovirus.
V dnešní době je už uznáván jako právoplatný nositel čtvrté domény.[zdroj?]
Mimivirus je rod virů s dvouvláknovou DNA, výjimečný svými rozměry (průměr dvacetistěnné kapsidy 400 nm, celkově až 800nm) i výjimečně velkým genomem (1 181 404 párů bází). Je znám jediný druh – Acanthamoeba polyphaga mimivirus (APMV). Virus byl nalezen v měňavkovci Acanthamoeba polyphaga a pro svou velikost byl zprvu považován za bakterii.
Mimivirus patří do skupiny tzv. jaderně-cytoplazmatických virů s velkou DNA označované NCLDV (z anglického nucleocytoplasmic large DNA viruses). Ta je někdy považována za novou, čtvrtou doménu živých organismů. Právě velký genom virů rodu mimivirus jako typických zástupců skupiny umožnil provést molekulárně biologické srovnání s ostatními doménami, poskytující indicie pro fylogenetické vyčlenění této domény. Existence nové domény je zatím přijímána skepticky.
Do příbuznosti mimiviru je řazen i rekordman ve velikosti virů z r. 2011 – Megavirus chilensis. Jeho genom je ještě o 6,5 % větší než genom mimiviru – obsahuje 1 259 197 párů bází. V roce 2013 pak rekord převzaly pandoraviry, pravděpodobně také z příbuznosti mimiviru – konkrétně Pandoravirus salinus obsahuje 2,47 milionů párů bází a s velikostí 1 mikrometr je dokonce viditelný optickým mikroskopem. V roce 2014 pozici rekordmana převzal ještě o polovinu větší pithovirus.
V dnešní době je už uznáván jako právoplatný nositel čtvrté domény.[zdroj?]
Mimivirus er en virusslægt der kun indeholder en kendt art, Acanthamoeba polyphaga mimivirus (APMV). Almindeligvis omtales APMV bare som "mimivirus". Den har det største capsid (proteinkapsel) af alle kendte virus, og et stort og kompleks genom i forhold til andre vira. Kendskabet til den er stadig ret begrænset; men opdagelsen af den begejstrede mange pga. de mulige egenskaber der kunne tænkes. Nogle anså den for et eksemplar i et helt nyt livsdomæne, mens andre anså den for et bindeled mellem vira og bakterier.
Mimivirus ist eine Gattung von Viren aus der Familie der Mimiviridae,[6] denen Amöben als natürliche Wirte dienen. Mit der Familie Mimiviridae gehört Mimivirus zu den Riesenviren im Phylum der Nucleocytoviricota (auch englisch Nucleocytoplasmic large DNA viruses, NCLDV; ursprünglicher Vorschlag „Nucleocytoplasmaviricota“; ein noch früherer Vorschlag ist der einer Ordnung „Megavirales“ s. l.[7] [5]).
In der Gattung Mimivirus gibt es mit Stand März 2019 eine einzige vom International Committee on Taxonomy of Viruses (ICTV) bestätigte Art Acanthamoeba polyphaga mimivirus (APMV), die daher auch die Typusart ist.[8] In der Umgangssprache und in älterer Literatur wird APMV üblicherweise nur als Mimivirus bezeichnet. Es gibt jedoch eine ganze Reihe phylogenetisch verwandter großer Viren, die als weitere Mitglieder dieser Gattung vorgeschlagen sind.
Das Mimivirus APMV wurde 1992 bei Forschungsarbeiten über die Legionärskrankheit (Legionellose) in einem Industriekühlturm in Bradford (England) entdeckt, wobei man feststellte, dass es sich in der Amöbe Acanthamoeba polyphaga vermehrt. Im Jahr 2003 wurde es an der Université de la Méditerranée in Marseille von einer Arbeitsgruppe um Didier Raoult identifiziert.[9] Mit einem Durchmesser von 400 nm haben die Viruspartikel (Virionen) von APMV die Größe von kleinen Bakterien.[10] Wegen dieser Größe und der äußeren Ähnlichkeit mit kugelförmigen Bakterien (Kokken) hielt man es zunächst für ein grampositives Bakterium und nannte es Bradfordcoccus. Als man den Irrtum erkannte, benannte man das neu entdeckte Virus in Anspielung auf seine Größe und Färbungseigenschaften Mimicking Virus, täuschendes Virus. Schließlich wurde daraus kurz Mimivirus, mit dem Namensteil mimi als eine Abkürzung für englisch mimicking microbe. Im Oktober 2004 wurde dann von Didier Raoult und Kollegen die Struktur seines Erbguts in der Fachzeitschrift Science veröffentlicht.[11] Mimivirus blieb übrigens nicht das einzige Virus, dem das passierte: Ein weiteres Beispiel ist das Misannotatedvirus (vorgeschlagen für Pithoviridae), zunächst Amina Cherif Louazani, Sarah Aherfi, Rania Francis, Rodrigo Rodrigues, Ludmila Santos Silva, Dehia Sahmi, Said Mougari, Nisrine Chelkha, Meriem Bekliz, Lorena Silva, Felipe Assis, Fábio Dornas, Jacques Yaacoub Bou Khalil, Isabelle Pagnier, Christelle Desnues, Anthony Levasseur, Philippe Colson, Jônatas Abrahão, Bernard La Scola.[12]
Das gleiche Team, das das APMV entdeckte, entdeckte später ein etwas größeres Virus, das Acanthamoeba castellanii mamavirus (ACMV, kurz Mamavirus) zusammen mit dem Sputnik-Virophagen, der es infiziert. ACMV und APMV sind so nahe verwandt, dass sie gewöhnlich in dieselbe Gattung Mimivirus gestellt werden.[13]
Bis 2013, als ein noch größeres Virus, das Pandoravirus, beschrieben wurde, wiesen die Viren der Gattung Mimivirus den größten Kapsiddurchmesser aller bekannten Viren auf; inzwischen wird es aber auch von nahen Verwandten Megavirus chilensis, Tupanvirus, Platanovirus, sowie anderen Riesenviren wie dem Pithovirus übertroffen.
Der erste bekannte Mimivirus-Wirt ist die Amöbe Acanthamoeba polyphaga (Gattung Acanthamoeba, Amoebozoa). Bisher konnten labormäßig nur Vertreter der Gattung Acanthamoeba, außer A. polyphaga noch A. castellanii und A. mauritaniensis, als Wirte dieses Virus eingesetzt werden, keine Zellen anderer einzelliger oder mehrzelliger Organismen.[14] Die natürlichen Wirte sind unbekannt (Stand 2015).[15]
Die Kapside der Virionen (Viruspartikel) von APMV erscheinen unter einem Elektronenmikroskop sechseckig, daher ist die Kapsidgeometrie ikosaedrisch.[17] Es scheint keine äußere virale Hülle zu geben, was darauf hindeutet, dass das Virus die Wirtszelle nicht durch Exozytose verlässt.[14]
Das Hauptprotein des Mimivirus-Kapsids besteht aus zwei Domänen vom Biskuitrollen-Typ (englisch Jelly roll fold). Dieses Protein bildet homotrimere Kapsomere als Organisationseinheit der Kapside. Die Kapsomere sind hexagonal in Form von „Gänseblümchen“ gepackt: Sechs Kapsomere umgeben eine Vertiefung zwischen ihnen.[17][18]
Die Virionen von Mimivirus haben einen Kapsiddurchmesser von 400 nm.
Das Kapsid ist mit einer kompakten Schicht von Fibrillen bedeckt (Tegument). Die aus der Oberfläche des Kapsids herausragenden Proteinfilamente (Fibrillen) haben eine Länge von etwa 100 nm (80–125 nm), und bringen damit die Gesamtlänge eines Virions auf 600 nm.[5][15] Abweichungen in der wissenschaftlichen Literatur lassen die Zahlen als sehr ungenau erscheinen, wenn zum Beispiel die ‚Größe’ des Virions gelegentlich als irgendwo zwischen 400 und 800 nm angegeben wird. Abgesehen von Unterschieden zwischen den einzelnen Virusstämmen in der Gattung Mimivirus ist manchmal die Gesamtgröße mit Filamenten, und manchmal der reine Kapsiddurchmesser angegeben.
Untersuchungen dieser Filamente von Klose et al. (2010) unter Verwendung eines Rasterkraftmikroskops ergeben, dass diese häufig an einer gemeinsamen Tragstruktur befestigt sind. Es konnte jedoch damals noch nicht herausgefunden werden, an welchen Teilen der Kapsidoberfläche diese Träger befestigt sind.[17] Jede Fibrille endet mit einer kleinen kugeligen Kappe aus einem Protein mit unbekannter Funktion.[15][17] Die Proteinfilamente erwiesen sich als resistent gegen Proteasen, außer sie wurden mit Lysozym behandelt. Die Filamente schienen deshalb mit Peptidoglycan beschichtet zu sein. Das war alles in guter Übereinstimmung mit der Tatsache, dass sich das Mimivirus durch die Gram-Methode anfärben lässt.[19]
Die Filamente spielen mit ihrer stark glykosylierten Oberfläche offenbar eine wichtige Rolle bei der Annäherung an die Wirtsamöben und der nachfolgenden Infektion.[19][15] Der Hauptbestandteil der Fasern ist das Protein R135 (neben L725 und L829). Seine Struktur ähnelt Proteinen aus der Familie der Glucose-Methanol-Cholin-Oxidoreduktasen (GMC-Oxidoreduktasen), die eine N-terminale FAD-Bindungsdomäne und eine C-terminale Substraterkennungsdomäne aufweisen. Das R135 am nächsten kommende Homolog ist eine Arylalkohol-Oxidase[20], die am biologischen Ligninabbau von Pflanzenzellwänden beteiligt ist. Somit könnte R135 an der Perforation der Zellwand ihrer natürlichen Wirte, insbesondere ligninhaltiger Algen, beteiligt sein.[15] Unter Laborbedingungen ist aber keines der drei genannten Proteine für die Infektiosität unbedingt erforderlich.[15]
Auffällig ist die fünfeckige, sternförmige Struktur an einer der Ecken des Kapsids, das so genannte ‚Stargate‘ (auch star-gate,[21] englisch für „Sternentor“, deutsch auch ‚Sternstruktur‘ oder ‚Seesternstruktur‘ genannt).[22] Blickt man direkt auf diesen Eckpunkt (den Mittelpunkt des Sterns), so scheinen zwischen dessen Strahlen fünf dreieckige Flächen zu liegen. Die Strahlen haben eine Breite von ungefähr 50 nm, eine Dicke von 40 nm und eine Länge von 200 nm; sie erreichen fast die benachbarten Eckpunkte der ikosaedrischen Kapsids. Das Stargate ist nicht von Fibrillen bedeckt.[15] Das Vorhandensein dieser Struktur verändert die Geometrie des Kapsids, indem seine Geometrie von der idealen Ikosaederform abweicht: Tatsächlich verläuft bei genauer Betrachtung nur eine einzige Achse mit Fünfstrahlsymmetrie durch das Virion, die durch den Mittelpunkt des Sterns (genannt Scheitelpunkt) verläuft.[17][23]
Die Symmetrie des Kapsids wird unterschiedlich angegeben mit einer Triangulationszahl T=972–1141 oder T=1200.[6]
Da auf der Oberfläche der Sternstruktur keine hexagonal geordneten Vertiefungen zu beobachten sind, wird vermutet, dass es sich bei dieser um ein Protein handelt, das sich vom Hauptkapsidprotein unterscheidet.[17]
Das Stargate spielt eine besondere Rolle bei der Infektion der Wirtszelle: Während der Infektion öffnet sich der „Verschluss“ am Scheitelpunkt und es erfolgt die Freisetzung des viralen Kerns (mit DNA und vorgefertigten Proteinen) aus dem Kapsid in das Zytosol der Wirtszelle (per Phagocytose). Das ist der Grund, warum die Sternstruktur als „Sternentor“ (englisch stargate) bezeichnet wird.[24]
Das Mimivirus hat mehrere morphologische Merkmale mit allen Mitgliedern der NCLDV-Virengruppe gemeinsam. Unmittelbar unter dem Kapsid des Mimivirus befinden sich beispielsweise zwei elektronendichte Schichten, die als Membranen gedeutet werden.[19] Unter diesen Membranen befindet sich eine etwa 7 nm dicke Proteinhülle, in der die lineare doppelsträngige DNA des Virus eingeschlossen ist. Dieser kondensierte Zentralkern des Virions erscheint unter dem Elektronenmikroskop als dunkler Bereich, das sogenannte ‚Nukleokapsid‘. In diesem Bereich befindet sich das große Genom des Virus, daneben auch mRNAs und vorgefertigte Proteine. Da alle anderen NCLDVs eine interne Lipidschicht besitzen, die den zentralen Kern umgibt, vermutet man das auch bei Mimivirus. Die Wände des Nukleokapsids liegen etwa 30 nm hinter den Wänden des Kapsids zurück, im Bereich der Sternstruktur (dem Stargate) ist die Oberfläche des Nukleokapsids zusätzlich abgesenkt.[17] Es wird angenommen, dass der Raum zwischen der Spitze der Sternstruktur und dem Nukleokapsid mit hydrolytischen Enzymen gefüllt ist, die für das Eindringen des Virus in die Wirtszelle erforderlich sind. Zwischen dem Kapsid und dem Nukleokapsid wurden interne Proteinstränge entdeckt, die anscheinend die gegenseitige räumliche Positionierung der beiden Teile zueinander stabilisieren.[19]
APMV hat damit im Vergleich zu den meisten anderen Viren ein großes und komplexes Genom, das aus einem einzelnen linearen DNA-Doppelstrang (dsDNA) besteht. Die Genomlänge des APMV Wildtyps (Ausgangsvariante Mimivirus M1) wurde von Raoult et al. (2004) mit 1.181.404 bp angegeben,[11] dieser Wert wurde durch Bäckström et al. (2019) leicht korrigiert auf 1.181.594 bp.[25] Das entspricht etwa 800 nm. Die fiberlose Variante Mimivirus M4 hat nur 0,993 Mbp, dazwischen liegen M2 mit 1.10 Mbp und M3 mit 1,10 Mbp.[26] Der GC-Gehalt von APMV liegt bei 28 %. Es gibt bei APMV 1260 Offene Leserahmen (ORFs, englisch open reading frames),[11] darunter vorhergesagt 979 kodierende Gene.[25][15][27] Dies geht weit über die Mindestausstattung von 4 Genen hinaus, die für ein Virus erforderlich sind, wie etwa bei den Phagen MS2 und Qβ.[28] Detaillierte Studien zum Genom korrigieren immer wieder Sequenzfehler und entdecken unter Umständen neue Leserahmen.[29]
Der Anteil von nichtcodierender DNA beträgt damit nur etwa 9,5 bis 10 %. Offene Leserahmen sind durch Lücken von ungefähr 157 Nukleotidpaaren getrennt. Zwei DNA-Abschnitte mit der Bezeichnung R (englisch right – rechts) und L (englisch left – links) kodieren ungefähr die gleiche Anzahl von Genen (450 bzw. 465, gemäß Daten von 2010). Der GC-Gehalt ist mit 28 % niedrig. In der Nähe der Enden des DNA-Moleküls wurden ‚Invertierte Wiederholungen‘ (englisch inverted repeats) mit 617 Nukleotidpaaren gefunden. Es wird vermutet, dass die gegenseitige Wechselwirkung dieser Stellen zur Bildung einer Q-Struktur führen kann – zirkuläre DNA mit zwei kleinen Fortsätzen.[29]
Bei der Analyse stieß man auf mindestens 21 Gene mit Homologie zu bekannten Proteinen, darunter solche, die man bis dato von keinem anderen Virus, sondern nur von zellulären Organismen kannte, inklusive Aminoacyl-tRNA-Synthetase.[11][9][30] 43 Gene sind homolog zu solchen anderer Riesenviren (NCLDVs).[15] Wie andere Riesenviren enthält Mimivirus mehrere Gene für den Zucker-, Lipid- und Aminosäurestoffwechsel. Es gab auch Stoffwechselgene, die zuvor in keinem anderen Virus gefunden wurden.[14]
Aus gereinigten Virionen konnten mehrere mRNA-Transkripte gewonnen werden. Wie schon bei anderen NCLDVs wurden insbesondere Transkripte für DNA-Polymerase, ein Kapsidprotein und ein TFII-ähnlicher Transkriptionsfaktor gefunden werden. Es wurden jedoch auch drei verschiedene Aminoacyl-tRNA-Synthetase-Transkripte und vier unbekannte mRNA-Moleküle gefunden, die für das Mimivirus spezifisch sind. Diese vorverpackten Transkripte können ohne virale Genexpression translatiert werden und sind wahrscheinlich für die Replikation von Mimivirus erforderlich. Andere DNA-Viren, wie das Humane Cytomegalievirus und das Herpes-simplex-Virus Typ 1, enthalten ebenfalls gepackte mRNA-Transkripte.[14]
Mimivirus ist eines der wenigen dsDNA-Viren, in deren Genom eine Intein-kodierende Sequenz nachgewiesen wurde. Inteine sind Proteindomäne, die ihre eigene Entfernung von einem Trägermolekül und die anschließende Verknüpfung der gebildeten Enden katalysieren. Eine solche Sequenz ist im Mimivirus-Gen für DNA-Polymerase B vorhanden.[31]
Auf Grund der außergewöhnlich komplexen genetischen Ausstattung des Virus stellt sich für einige Forscher die Frage neu, wo die Grenze zwischen belebter und unbelebter Natur verlaufe, also wie „Lebewesen“ zu definieren ist.
Die Einzelheiten und die verschiedenen Stadien im Replikationszyklus von Mimivirus, wie die offensichtliche Bindung an die Zelloberfläche und den Eintritt in die Zelle, die Freisetzung des Viruskerns, die DNA-Replikation, die Transkription, die Translation, und schließlich den Zusammenbau und die Freisetzung von Tochter-Virionen, sind noch nicht ausreichend bekannt. Die Wissenschaftler haben jedoch den oben angegebenen allgemeinen Überblick anhand elektronenmikroskopischer Aufnahmen infizierter Zellen erstellt. Alle Stadien des Vermehrungszyklus verlaufen im Cytoplasma der Wirtszelle.[32]
Die Infektion der Amöbe mit einem Mimivirus erfolgt vermutlich nach folgendem Szenario:
Die Übertragung geschieht durch passive Diffusion.[6]
Es wurde spekuliert, dass Mimivirus Erreger bestimmter Formen von Lungenentzündung (Pneumonia) sein könnte. Dies beruht hauptsächlich auf indirekten Nachweisen in Form von Antikörpern gegen das bei Lungenentzündungspatienten entdeckte Virus.[33] Aufgrund der wenigen bisherigen Veröffentlichungen ist die Einstufung des Mimivirus als möglicher Krankheitserreger derzeit jedoch schwierig. Ein großer Teil der Fälle von Lungenentzündung verläuft ohne feststellbare Ursache.[34] Zwar wurde ein Mimivirus bei einer an Lungenentzündung leidenden Tunesierin isoliert,[35] und es gibt aus Zellkulturen Hinweise darauf, dass Mimivirus Makrophagen infizieren kann und darin repliziert wird.[36] So wurde unter experimentellen Bedingungen beobachtet, dass Mimivirus humane Makrophagen infizieren kann, d. h. via Phagocytose in die Zellen eindringen, und sich dort replizieren kann.[37][36] Außerdem wurden in mehreren Studien bei einer kleinen Anzahl von Patienten mit Lungenentzündung Antikörper gegen das Mimivirus gefunden.[33][38] Es wurde auch ein Einzelfall einer Lungenentzündung eines Laborassistenten beschrieben, der mit Kulturen dieses Virus arbeitete. Der Gehalt an Antikörpern gegen Mimivirus in seinem Blut war ebenfalls erhöht.[39] Das Vorhandensein von Antikörpern gegen das Virus an sich ist jedoch kein Hinweis auf seine Pathogenität. Es ist möglich, dass das Mimivirus einfach starke immunogene Eigenschaften aufweist, d. h. eine deutliche Immunantwort auslöst.[29] Auch war es in keinem der registrierten Fälle möglich, das Virus in seiner reinen Form aus Proben von Flüssigkeiten zu isolieren, die von Patienten erhalten wurden.[40]
Die Fiberproteine R135 und L829 wurden als Hauptantigene des Mimivirus identifiziert: Die faserlose Variante Mimivirus M4 zeigte jedoch keine Reaktivität mit Seren von menschlichen Patienten, was bestätigt, dass diese Proteine in M4 fehlen.[15]
Das Zamilon-Virus ist ein Satellitenvirus, das Mimiviren der Linien B und C befällt, nicht aber die Mimiviren der Linie A. Diese weisen nämlich eine MIMIVIRE, englisch mimivirus virophage resistance element, genannte Resistenz auf, die ähnlich funktioniert wie das CRISPR/Cas-System.[41][42]
Im Übrigen wurde nachgewiesen, dass es nicht nur einen Gentransfer zwischen den amöboid Wirten und Riesenviren als intrazellulären viralen Endocytobionten (Organismen, die in den Zellen anderer Organismen leben oder sich vermehren) gibt[43] sondern sogar zwischen den Viren und gleichzeitig vorhandenen bakteriellen Endocytobionten.[44][26][45]
Die Gattung Mimivirus und einige andere – vom ICTV mit Stand März 2019 noch nicht bestätigte – genetisch ähnliche Gattungen und Spezies der Familie Mimiviridae (wie zum Beispiel „Mamavirus“, „Megavirus“ und „Moumouvirus“) bilden eine als Gruppe I bezeichnete Klade der Mimiviren im weiteren Sinn. Für diese wurde vorgeschlagen, sie als „Megamimivirinae“,[3] „Megavirinae“[2] oder auch „Mimivirinae“[1] in den Rang einer Unterfamilie zu erheben und so von anderen ebenfalls vorgeschlagenen Unterfamilien der Mimiviridae abzugrenzen, nämlich:
Innerhalb der Mimiviridae-Gruppe I zeichnen sich die folgenden Linien ab:[2]
Da bisher (März 2019) vom ICTV lediglich die Gattung Mimivirus bestätigt wurde, bleibt zurzeit noch offen, welche Kandidaten für diese vorgeschlagene Unterfamilie dieser Gattung einmal zugeordnet werden, und für welche eigene Gattungen einzurichten sind. Sicher scheint aber, dass die Vertreter der Linie A in jedem Fall zur Gattung Mimivirus gehören.
Der von Wilson et al. 2017 beschriebene Kandidat „gvSAG AB-566-O17“ (vom NCBI als Spezies „Mimivirus AB-566-O17“ bezeichnet[47]) ist nach den Autoren (Fig. 2) mit APMV weitläufiger verwandt als CroV (aber näher als Vertreter der OLPG-Gruppe). Er ist daher keiner der obigen Gruppen zuzuordnen (insbesondere nicht der Gattung Mimivirus).[48]
Systematik der Linie A (Mimiviren im engeren Sinn):
Vorgeschlagener phylogenetischer Baum der Gattung Mimivirus nach Abrahão et al. (2018), Fig. 4:[107]
Mimivirus Mimivirus-Gruppe A„Acanthamoeba castellanii mamavirus“ (ACMV)
Hirudovirus
Acanthamoeba polyphaga mimivirus (APMV) - Wildtyp M1;
Mimivirus shirakomae, Mimivirus kasaii
Kroon-Virus
„Terra2-Virus“
„Lentille-Virus“
„Niemeyer-Virus“
Mimivirus Bombay
Sambavirus
Mimivirus-Gruppe B (Moumouvirus-Gruppe)
Mimivirus-Gruppe C (Courdo11-Gruppe: Megavirus chilensis,…)
Anmerkung: Kursivschrift für vorgeschlagene Spezies.
Mimivirus ist eine Gattung von Viren aus der Familie der Mimiviridae, denen Amöben als natürliche Wirte dienen. Mit der Familie Mimiviridae gehört Mimivirus zu den Riesenviren im Phylum der Nucleocytoviricota (auch englisch Nucleocytoplasmic large DNA viruses, NCLDV; ursprünglicher Vorschlag „Nucleocytoplasmaviricota“; ein noch früherer Vorschlag ist der einer Ordnung „Megavirales“ s. l. ).
In der Gattung Mimivirus gibt es mit Stand März 2019 eine einzige vom International Committee on Taxonomy of Viruses (ICTV) bestätigte Art Acanthamoeba polyphaga mimivirus (APMV), die daher auch die Typusart ist. In der Umgangssprache und in älterer Literatur wird APMV üblicherweise nur als Mimivirus bezeichnet. Es gibt jedoch eine ganze Reihe phylogenetisch verwandter großer Viren, die als weitere Mitglieder dieser Gattung vorgeschlagen sind.
Mimivirus is a genus of giant viruses, in the family Mimiviridae. Amoeba serve as their natural hosts.[2][3] This genus contains a single identified species named Acanthamoeba polyphaga mimivirus (APMV). It also refers to a group of phylogenetically related large viruses.[4]
In colloquial speech, APMV is more commonly referred to as just "mimivirus". Mimivirus, short for "mimicking microbe", is so called to reflect its large size and apparent Gram-staining properties.[5]
Mimivirus has a large and complex genome compared with most other viruses. Until 2013, when a larger virus Pandoravirus was described, it had the largest capsid diameter of all known viruses.[6]
APMV was discovered accidentally in 1992 within the amoeba Acanthamoeba polyphaga, after which it is named, during research into legionellosis by researchers from Marseille and Leeds.[7] The virus was observed in a Gram stain and mistakenly thought to be a Gram-positive bacterium. As a consequence it was named Bradfordcoccus, after Bradford, England, where the amoeba had originated. In 2003, researchers at the Université de la Méditerranée in Marseille, France, published a paper in Science identifying the micro-organism as a virus. It was given the name "mimivirus" (for "mimicking microbe") as it resembles a bacterium on Gram staining.[8]
The same team that discovered the mimivirus later discovered a slightly larger virus, dubbed the mamavirus, and the Sputnik virophage that infects it.[9]
Mimivirus has been placed into a viral family by the International Committee on Taxonomy of Viruses as a member of the Mimiviridae,[10] and has been placed into Group I of the Baltimore classification system.[11]
Although not strictly a method of classification, mimivirus joins a group of large viruses known as nucleocytoplasmic large DNA viruses (NCLDV). They are all large viruses which share both molecular characteristics and large genomes. The mimivirus genome also possesses 21 genes encoding homologs to proteins which are seen to be highly conserved in the majority of NCLDVs, and further work suggests that mimivirus is an early divergent of the general NCLDV group.[8]
The mimivirus is the fourth-largest virus, followed by the Megavirus chilensis, Pandoravirus and Pithovirus. Mimivirus has a capsid diameter of 400 nm. Protein filaments measuring 100 nm project from the surface of the capsid, bringing the total length of the virus up to 600 nm. Variation in scientific literature renders these figures as highly approximate, with the "size" of the virion being casually listed as anywhere between 400 nm and 800 nm, depending on whether total length or capsid diameter is actually quoted.
Its capsid appears hexagonal under an electron microscope, therefore the capsid symmetry is icosahedral.[12] It does not appear to possess an outer viral envelope, suggesting that the virus does not exit the host cell by exocytosis.[13] Mimivirus shares several morphological characteristics with all members of the NCLDV group of viruses. The condensed central core of the virion appears as a dark region under the electron microscope. The large genome of the virus resides within this area. An internal lipid layer surrounding the central core is present in all other NCLDV viruses, so this features may also be present in mimivirus.[12]
Several mRNA transcripts can be recovered from purified virions. Like other NCLDVs, transcripts for DNA polymerase, a capsid protein and a TFII-like transcription factor were found. However, three distinct aminoacyl tRNA synthetase enzyme transcripts and four unknown mRNA molecules specific to mimivirus were also found. These pre-packaged transcripts can be translated without viral gene expression and are likely to be necessary to Mimivirus for replication. Other DNA viruses, such as the Human cytomegalovirus and Herpes simplex virus type-1, also feature pre-packaged mRNA transcripts.[13]
The mimivirus genome is a linear, double-stranded molecule of DNA with 1,181,404 base pairs in length.[14] This makes it one of the largest viral genomes known, outstripping the next-largest virus genome of the Cafeteria roenbergensis virus by about 450,000 base pairs. In addition, it is larger than at least 30 cellular clades.[15]
In addition to the large size of the genome, mimivirus possesses an estimated 979 protein-coding genes, far exceeding the minimum 4 genes required for viruses to exist (c.f. MS2 and Qβ viruses).[16] Analysis of its genome revealed the presence of genes not seen in any other viruses, including aminoacyl tRNA synthetases, and other genes previously thought only to be encoded by cellular organisms. Like other large DNA viruses, mimivirus contains several genes for sugar, lipid and amino acid metabolism, as well as some metabolic genes not found in any other virus.[13] Roughly 90% of the genome was of coding capacity, with the other 10% being "junk DNA".

The stages of mimivirus replication are not well known, but as a minimum it is known that mimivirus attaches to a chemical receptor on the surface of an amoeba cell and is taken into the cell. Once inside, an eclipse phase begins, in which the virus disappears and all appears normal within the cell. After about 4 hours small accumulations can be seen in areas of the cell. 8 hours after infection many mimivirus virions are clearly visible within the cell. The cell cytoplasm continues to fill with newly synthesised virions, and about 24 hours after initial infection the cell likely bursts open to release the new mimivirus virions.[13]
Little is known about the details of this replication cycle, most obviously attachment to the cell surface and entry, viral core release, DNA replication, transcription, translation, assembly and release of progeny virions. However, scientists have established the general overview given above using electron micrographs of infected cells. These micrographs show mimivirus capsid assembly in the nucleus, acquisition of an inner lipid membrane via budding from the nucleus, and particles similar to those found in many other viruses, including all NCLDV members. These particles are known in other viruses as viral factories and allow efficient viral assembly by modifying large areas of the host cell.
Mimivirus may be a causative agent of some forms of pneumonia; this is based mainly on indirect evidence in the form of antibodies to the virus discovered in pneumonia patients.[17] However, the classification of mimivirus as a pathogen is tenuous at present as there have been only a couple of papers published potentially linking mimivirus to actual cases of pneumonia. A significant fraction of pneumonia cases are of unknown cause,[18] though a mimivirus has been isolated from a Tunisian woman suffering from pneumonia.[19] There is evidence that mimivirus can infect macrophages.[20]
Mimivirus shows many characteristics which place it at the boundary between living and non-living. It is as large as several bacterial species, such as Rickettsia conorii and Tropheryma whipplei, possesses a genomic size comparable to that of several bacteria, including those above, and codes for products previously not thought to be encoded by viruses (including a kind of collagen[21]). In addition, mimivirus has genes coding for nucleotide and amino acid synthesis, which even some small obligate intracellular bacteria lack. They do, however, lack any genes for ribosomal proteins, making mimivirus dependent on a host cell for protein translation and energy metabolism.[21]
Because its lineage is very old and could have emerged prior to cellular organisms,[22][23] Mimivirus has added to the debate over the origins of life. Some genes that code for characteristics unique to Mimivirus, including those coding for the capsid, have been conserved in a variety of viruses which infect organisms from all domains. This has been used to suggest that Mimivirus is related to a type of DNA virus that emerged before cellular organisms and played a key role in the development of all life on Earth.[22] An alternative hypothesis is that there were three distinct types of DNA viruses that were involved in generating the three known domains of life—eukarya, archaea and bacteria.[23] It has been suggested that Mimivirus and similar kinds are remnants of a "fourth domain" of life, and that other giant virus may represent other ancient domains.[21]
Nevertheless, mimivirus does not exhibit the following characteristics, all of which are part of many conventional definitions of life:
 Schematic drawing of a virion of genus Mimivirus (cross section and side view) showing filaments ("hairs") and stargate (downside).
Schematic drawing of a virion of genus Mimivirus (cross section and side view) showing filaments ("hairs") and stargate (downside). Mimivirus is a genus of giant viruses, in the family Mimiviridae. Amoeba serve as their natural hosts. This genus contains a single identified species named Acanthamoeba polyphaga mimivirus (APMV). It also refers to a group of phylogenetically related large viruses.
In colloquial speech, APMV is more commonly referred to as just "mimivirus". Mimivirus, short for "mimicking microbe", is so called to reflect its large size and apparent Gram-staining properties.
Mimivirus has a large and complex genome compared with most other viruses. Until 2013, when a larger virus Pandoravirus was described, it had the largest capsid diameter of all known viruses.
Mimivirus es un género de virus de gran tamaño que infecta amebas. Poseen ADN de doble cadena cuyas partículas maduras tienen una cápsida de entre 400 y 500 nm de diámetro, 1 181 404 pb y aproximadamente 911 ORF.[1] Fue descubierto por primera vez en 1992 en una torre de refrigeración industrial en Bradford (Inglaterra) e identificado en 2003 por un grupo de investigadores en la Universidad de Aix-Marsella (Francia).[2] El virus, que fue descubierto mientras se estudiaba la legionelosis (una enfermedad causada por la bacteria Legionella), se encontró dentro de la ameba acuática Acanthamoeba polyphaga. A su vez, también se encontraron anticuerpos para este virus en muestras de sangre humana.[3]
Debe su nombre a que cuando fue descubierto fue confundido con una bacteria debido a su tamaño, y a que "imitaba" (en inglés mimic) mediante una tinción de Gram la tinción de una bacteria Gram positiva. Los Mimivirus son los únicos miembros de una nueva familia de virus, denominada Mimiviridae, que se integra dentro de un grupo de virus de origen monofilético, denominado virus nucleocitoplasmáticos de ADN de gran tamaño (NCLDV, en inglés) y al que pertenecen virus tan relevantes para la salud humana y animal como el virus de la viruela (Familia Poxviridae), el virus del herpes labial (Herpesviridae) o el virus de la peste porcina africana o VPPA (Asfarviridae).
Se realizaron estudios en la Université de la Méditerranée y publicados en la revista Nature en los que se secuenció el genoma del organismo.[4] Los resultados obtenidos fueron: 800 nm de longitud, 1 181 404 pares de bases, y 1262 genes. Solo el 10 % del genoma es ADN basura. Jean-Michel Claverie, de esta universidad, ha dicho acerca del Mimi: "Hace parecer a este tipo de virus a ADN como una nueva forma de vida parasitaria".[2]
Actualmente se está investigando sobre la posibilidad de que la presencia de Mimivirus en pacientes con cuadros graves de enfermedad pulmonar esté asociada a un pronóstico negativo, no obstante se han publicado datos contradictorios en cuanto a esto.[5][6][7] En cualquier caso tanto por su peculiaridad como por su asociación a microorganismos causantes de graves neumonías infecciosas, se trata de un organismo modelo de gran interés científico.
Varios miembros de la comunidad científica han declarado recientemente que, debido a que la partícula viral es capaz de generar sus propias proteínas, es de hecho un organismo vivo, una idea que no hace más que agudizar la confusión existente con respecto a la clasificación de los virus. Mimi, con sus 911 genes codificantes, codifica 50 proteínas que nunca antes habían sido vistas en un virus, como un citocromo P450, implicado en el metabolismo energético. Además codifica proteínas que llevan mecanismos bioquímicos que sí tienen algunos virus a una complejidad máxima, como son chaperonas que asisten en el correcto plegamiento de las proteínas y enzimas que pueden proporcionar mecanismos para mantener la integridad del ADN (rutas bioquímicas de reparación de errores durante la replicación o de daños físico-químicos). Representa una nueva familia dentro de los grandes virus ADN nucleocitoplasmáticos que emergieron en la Tierra hace aproximadamente cuatro mil millones de años. Además, desafía el sistema de clasificación de los organismos dentro de tres dominios distintos (Archaea, Bacteria y Eukarya), ya que parece exigir la creación de un cuarto dominio. Su linaje es muy antiguo y podría haber emergido antes de la existencia de los organismos celulares, lo cual puede traer consecuencias drásticas en las concepciones sobre el origen de la vida.
El genoma de Mimivirus está conformado por una molécula de ADN de doble cadena lineal de 1 181 404 pares de bases. Esto hace que sea el mayor genoma viral conocido, superando al del miovirus Bacillus phage G. Además del gran tamaño de su genoma, se estima que Mimivirus posee 911 genes que codifican proteínas, superando considerablemente los 4 genes codificantes mínimos requeridos para la existencia de los virus (véase MSD y virus Qβ).
El análisis del genoma de Mimivirus mostró la presencia de genes no encontrados en ningún otro virus. Un ejemplo es el gen que codifica para ARNt aminoacil transferasa, que hasta entonces se creía exclusivo de organismos celulares. Como otros virus de gran tamaño, Mimivirus contiene varios genes para el metabolismo de azúcares, lípidos y aminoácidos, así como otros genes relacionados con el metabolismo y no encontrados en ningún otro virus. Aproximadamente el 90 % del genoma es codificante, siendo el 10 % restante "ADN basura".
El Mimivirus posee un gran número de características que lo sitúan en la frontera entre lo vivo y lo no vivo. Es tan grande como algunas especies de bacterias, tales como Rickettsia conorii y Tropheryma whipplei, el tamaño de su genoma es comparable al de varias bacterias, incluidas las anteriormente nombradas, y codifica para productos que se pensaba que no eran codificados por ningún virus. Además, Mimivirus posee genes que codifican para la síntesis de nucleótidos y aminoácidos, algunos de los cuales están ausentes incluso en pequeños procariotas intracelulares obligados. Esto significa, que al contrario que estas bacterias, Mimivirus no depende del genoma de la célula hospedadora para codificar las rutas metabólicas para estos productos. Sin embargo, Mimivirus carece de genes para proteínas ribosómicas, por lo que depende de la célula hospedadora para la traducción proteica y el metabolismo energético. Todos estos factores, en conjunto, han causado un debate científico sobre si el Mimivirus podría ser una forma de vida distinta, comparable a los dominios Eukarya, Archaea y Bacteria. Sin embargo, Mimivirus no posee las siguientes características, las cuales forman parte de alguno de los convenios sobre la definición de vida: homeostasis, crecimiento en el sentido normal del término (en lugar de replicarse a través del autoensamblaje de componentes individuales) o llevar a cabo la división celular.
Debido a que se piensa que podría haber existido con anterioridad a los organismos celulares, Mimivirus tiene un papel importante en el debate sobre los orígenes de la vida. Algunos genes exclusivos de Mimivirus, incluyendo aquellos que codifican para su cápsida, se han conservado en una gran variedad de virus que infectan organismos de todos los dominios —Eukarya, Archaea y Bacteria—. Esto hace pensar que Mimivirus está relacionado con un tipo de virus de ADN que existió antes que los organismos celulares y jugó un papel esencial en el desarrollo de la vida en la Tierra.[8] Una hipótesis alternativa afirma que fueron tres tipos distintos de virus de ADN los que dieron lugar a los tres dominios conocidos.[9]
|número-autores= (ayuda)
|número-autores= (ayuda)
Mimivirus es un género de virus de gran tamaño que infecta amebas. Poseen ADN de doble cadena cuyas partículas maduras tienen una cápsida de entre 400 y 500 nm de diámetro, 1 181 404 pb y aproximadamente 911 ORF. Fue descubierto por primera vez en 1992 en una torre de refrigeración industrial en Bradford (Inglaterra) e identificado en 2003 por un grupo de investigadores en la Universidad de Aix-Marsella (Francia). El virus, que fue descubierto mientras se estudiaba la legionelosis (una enfermedad causada por la bacteria Legionella), se encontró dentro de la ameba acuática Acanthamoeba polyphaga. A su vez, también se encontraron anticuerpos para este virus en muestras de sangre humana.
Debe su nombre a que cuando fue descubierto fue confundido con una bacteria debido a su tamaño, y a que "imitaba" (en inglés mimic) mediante una tinción de Gram la tinción de una bacteria Gram positiva. Los Mimivirus son los únicos miembros de una nueva familia de virus, denominada Mimiviridae, que se integra dentro de un grupo de virus de origen monofilético, denominado virus nucleocitoplasmáticos de ADN de gran tamaño (NCLDV, en inglés) y al que pertenecen virus tan relevantes para la salud humana y animal como el virus de la viruela (Familia Poxviridae), el virus del herpes labial (Herpesviridae) o el virus de la peste porcina africana o VPPA (Asfarviridae).
Se realizaron estudios en la Université de la Méditerranée y publicados en la revista Nature en los que se secuenció el genoma del organismo. Los resultados obtenidos fueron: 800 nm de longitud, 1 181 404 pares de bases, y 1262 genes. Solo el 10 % del genoma es ADN basura. Jean-Michel Claverie, de esta universidad, ha dicho acerca del Mimi: "Hace parecer a este tipo de virus a ADN como una nueva forma de vida parasitaria".
Actualmente se está investigando sobre la posibilidad de que la presencia de Mimivirus en pacientes con cuadros graves de enfermedad pulmonar esté asociada a un pronóstico negativo, no obstante se han publicado datos contradictorios en cuanto a esto. En cualquier caso tanto por su peculiaridad como por su asociación a microorganismos causantes de graves neumonías infecciosas, se trata de un organismo modelo de gran interés científico.
Mimivirus on alun perin ameeboista löytynyt jättiläisvirus, joka muistuttaa bakteereita, mutta ei sisällä itsenäiseen lisääntymiseen tarvittavia soluelimiä eli ribosomeja. Se on poikkeuksellisen suuri, pyöreä ja halkaisijaltaan 750 nanometriä, kun virukseksi kookas isorokkovirus on vain 300 nanometriä ja poliovirus vain 30 nanometriä. Sillä on yli 900 geeniä ja yli miljoona emäsparia.[1]
Suuri virus sai nimen mimivirus, koska se jäljittelee bakteereja. Yleensä virukset eivät tarvitse paljon geenejä, koska ne kaappaavat isäntäsolun geneettisen koneiston omiin tarkoituksiinsa. Jättiläisviruksessa on muista viruksista poiketen useita geenejä, joita tarvitaan kopioitumiseen. Lisäksi on geenejä, jotka osallistuvat proteiinien laskostamiseen, korjaavat dna-vaurioita sekä auttavat rakentamaan ja kuljettamaan aminohappoja. Silti ne eivät pysty lisääntymään itsenäisesti, vaan ovat aitotumallisista riippuvaisia loisia. Jättiläisviruksia on kuitenkin epäilty solujen tuman esimuodoiksi.[1]
Mimiviruksen löytäminen liittyi Bradfordin kaupungin keuhkokuume-epidemiaan vuodelta 1992, jolloin niitä oli vesitornin ameeboissa. Jättiläisviruksia onkin löydetty ihmisistä usein keuhkokuumeiden yhteydessä, vaikka niitä pidetäänkin yleisesti ihmisille vaarattomina. Jättiläisvirukset tuottavat kollageenia, joka muistuttaa ihmisten omaa sidekudosproteiinia. Tutkijat epäilevätkin, että Nivelreumassa virukset laukaisevat hyökkäyksen ihmisen omaa sidekudosta vastaan. Testeissä mimiviruksen on todettu lisäävän hiirten riskiä sairastua reumaan. Mimiviruksen vasta-aineet ovatkin nivelreumaa sairastavilla terveitä yleisempiä. Mimivirus oli ensimmäinen löydetty ns. jättiläisvirus ja sen jälkeen on löydetty useita muita.[1]
Mimivirus on alun perin ameeboista löytynyt jättiläisvirus, joka muistuttaa bakteereita, mutta ei sisällä itsenäiseen lisääntymiseen tarvittavia soluelimiä eli ribosomeja. Se on poikkeuksellisen suuri, pyöreä ja halkaisijaltaan 750 nanometriä, kun virukseksi kookas isorokkovirus on vain 300 nanometriä ja poliovirus vain 30 nanometriä. Sillä on yli 900 geeniä ja yli miljoona emäsparia.
Suuri virus sai nimen mimivirus, koska se jäljittelee bakteereja. Yleensä virukset eivät tarvitse paljon geenejä, koska ne kaappaavat isäntäsolun geneettisen koneiston omiin tarkoituksiinsa. Jättiläisviruksessa on muista viruksista poiketen useita geenejä, joita tarvitaan kopioitumiseen. Lisäksi on geenejä, jotka osallistuvat proteiinien laskostamiseen, korjaavat dna-vaurioita sekä auttavat rakentamaan ja kuljettamaan aminohappoja. Silti ne eivät pysty lisääntymään itsenäisesti, vaan ovat aitotumallisista riippuvaisia loisia. Jättiläisviruksia on kuitenkin epäilty solujen tuman esimuodoiksi.
Mimiviruksen löytäminen liittyi Bradfordin kaupungin keuhkokuume-epidemiaan vuodelta 1992, jolloin niitä oli vesitornin ameeboissa. Jättiläisviruksia onkin löydetty ihmisistä usein keuhkokuumeiden yhteydessä, vaikka niitä pidetäänkin yleisesti ihmisille vaarattomina. Jättiläisvirukset tuottavat kollageenia, joka muistuttaa ihmisten omaa sidekudosproteiinia. Tutkijat epäilevätkin, että Nivelreumassa virukset laukaisevat hyökkäyksen ihmisen omaa sidekudosta vastaan. Testeissä mimiviruksen on todettu lisäävän hiirten riskiä sairastua reumaan. Mimiviruksen vasta-aineet ovatkin nivelreumaa sairastavilla terveitä yleisempiä. Mimivirus oli ensimmäinen löydetty ns. jättiläisvirus ja sen jälkeen on löydetty useita muita.
Mimivirus est un genre de virus à ADN. Les virus de ce taxon sont particulièrement grands, souvent plus gros que bien des bactéries, et de ce fait on les qualifie de girus — techniquement ils en sont le holotype. Au niveau écologique, Mimivirus infecte une amibe : Acanthamoeba polyphaga.
En 2009, son origine est sujette à discussion parmi les biologistes et certains voient en lui le représentant d'une nouvelle branche de l'arbre phylogénétique[1], donnant ainsi un argument supplémentaire en faveur des théories selon lesquelles les virus devraient être considérés comme des êtres vivants à part entière[2].
Jusqu'en 2012, Mimivirus était le quatrième plus grand virus connu — les premiers étant le virus ebola (dont la longueur peut dépasser 1 000 nm), Mamavirus[3], et Megavirus chilensis[4]. En 2013, une équipe de chercheurs marseillais découvre les Pandoravirus, soit deux virus encore plus gros et génétiquement plus complexes que certaines bactéries, dans des sédiments en mer au Chili, et en eau douce en Australie[5].
Parce qu'anciennement les biologistes et virologues n'imaginaient pas qu'un virus puisse surclasser en dimensions une bactérie, la découverte de la nature virale d'un parasite de Acanthamoeba polyphaga allait prendre plus de dix ans.
C'est en 1992 qu'est prélevée l'eau d'une tour de climatisation industrielle à Bradford, en Angleterre. Y sont identifiés Acanthamoeba polyphaga et la découverte d'un parasite nouveau : d'abord assimilé à une bactérie, les « cellules » sont dénommées Bradford coccus.
Ce n'est qu'en 2003 que sera identifié Mimivirus en tant que virus à l'université de la Méditerranée à Marseille[6].
La séquence complète de son génome a été publiée en 2004[7].
La description taxonomique est publiée en 2005 sous l'égide de l'Union Internationale des Sociétés Microbiologiques et de sa Division Virologie[8].
Officiellement, le professeur de microbiologie Didier Raoult a baptisé ce microbe du nom de Mimivirus parce que cela signifie « Mimicking Microbe Virus ».
Officieusement, ce serait en souvenir des aventures de « Mimi l'amibe », un héros de son enfance sorti de l'imagination de son père qui lui racontait de la sorte l'histoire de l'évolution[9].
À côté de toutes ses spécificités, Mimivirus tend à confirmer certains aspects de la définition d'un virus donnée par André Lwoff, notamment l'absence de ribosome et de ses protéines et l'incapacité de se diviser[10]. Son mode de réplication passe par une phagocytose du virion, puis après une phase d'éclipse la mise en place d'usine à virions.
De forme icosaédrique, il mesure 400 nanomètres de diamètre. Le matériel viral est introduit à son apex avant terminaison de la synthèse de la capside, alors que le vertex en forme d'étoile à cinq branches fusionnera avec la membrane du phagosome pour relarguer son contenu dans le cytoplasme hôte.
En plus de son ADN, il contient également de l'ARN et partage ainsi cette particularité avec les cytomégalovirus, mettant à mal ce dogme dans la définition des virus[10].
Le génome contient environ 1 181 404 paires de bases et 1 262 gènes[11]. C'était deux fois plus gros que les plus imposants virus identifiés à l'époque, plus gros que celui des plus petites bactéries[1].
Chose étrange, certains de ces gènes (une trentaine) ne sont pas présents chez les autres virus, mais le sont dans les organismes cellulaires, comme ceux codant des protéines de réparation de l'ADN ou de la traduction de l'ARN en protéines. Ces gènes ne devraient pas servir à Mimivirus, car les virus utilisent la machinerie cellulaire de leur hôte[10]. À tel point que certains ont relié Mimivirus à l'arbre de la vie aux côtés des trois domaines du vivant que sont les archées, les bactéries et les eucaryotes[1].
Ces affinités avec le vivant ne font pas pour autant de lui un virus vraiment à part comparé aux autres virus, car il partage des dizaines de gènes spécifiques aux virus et neuf gènes qui sont communs à l'ensemble des autres virus[10].
Son rôle comme agent de la pneumonie chez l'humain a été rapporté après la contamination d'un technicien de laboratoire[12]. Un modèle expérimental a montré qu'il pouvait provoquer des pneumonies chez la souris.
L'étude de Mimivirus par l'équipe de Didier Raoult a permis de découvrir en août 2008 un nouveau type de virus, surnommé Spoutnik. Ce dernier a la particularité de ne pas pouvoir infecter de cellule, ce qui le rend inapte à se multiplier par la seule méthode alors connue chez les virus. Pour accomplir son cycle de réplication, Spoutnik infecte une forme particulièrement grosse de Mimivirus, désignée sous le nom de Mamavirus, et détourne la machinerie cellulaire de la cellule-hôte, l'amibe Acanthamoeba polyphaga, que le Mimivirus a lui-même détournée. Ce nouveau type de virus parasitant d'autres virus a été dénommé virophage par analogie avec le terme bactériophage désignant les virus infectant les bactéries. Cette découverte est une première et démontre la richesse et la complexité de ces particules aux frontières du vivant[13].
Ce virus n'est en 2008 pas classé par l'International Committee on Taxonomy of Viruses (ICTV)[14]. Mais cette non-classification n'empêche cependant pas de le classer dans celle de Baltimore, où sa structure nucléique le range dans le groupe I des virus à ADN bicaténaires.
Par sa structure et la conservation de certains gènes spécifiques aux grands virus nucléocytoplasmiques (NCLDV), le mimivirus est classé dans ce groupe, mais il n'a cependant d'affinité particulière avec aucune des cinq familles de ce groupe, ce qui pousse certains virologistes à le classer au sein d'une nouvelle famille, les Mimiviridae[1].
Mimivirus est un genre de virus à ADN. Les virus de ce taxon sont particulièrement grands, souvent plus gros que bien des bactéries, et de ce fait on les qualifie de girus — techniquement ils en sont le holotype. Au niveau écologique, Mimivirus infecte une amibe : Acanthamoeba polyphaga.
En 2009, son origine est sujette à discussion parmi les biologistes et certains voient en lui le représentant d'une nouvelle branche de l'arbre phylogénétique, donnant ainsi un argument supplémentaire en faveur des théories selon lesquelles les virus devraient être considérés comme des êtres vivants à part entière.
Jusqu'en 2012, Mimivirus était le quatrième plus grand virus connu — les premiers étant le virus ebola (dont la longueur peut dépasser 1 000 nm), Mamavirus, et Megavirus chilensis. En 2013, une équipe de chercheurs marseillais découvre les Pandoravirus, soit deux virus encore plus gros et génétiquement plus complexes que certaines bactéries, dans des sédiments en mer au Chili, et en eau douce en Australie.
Os Mimivirus son un grupo de virus de gran tamaño que posúen ADN de cadea dupla e cuxas partículas maduras miden 400 nm de diámetro. O seu ADN posúe 800.000 pares de bases e 900 xenes.
O primeiro foi descuberto en 1992 nunha torre de refrixeración industrial en Bradford (Inglaterra) e identificado en 2003 por un grupo de investigadores da Université de le Méditerranée (Universidade do Mediterráneo) en Marsella (Francia). O virus, que foi descuberto mentres se estudaba a lexionelose (enfermiadade causada poa bacteria Legionella), encontrouse dentro da ameba acuática Acanthamoeba polyphaga. Á súa vez, tamén se encontraron anticorpos para este virus en mostras de sangue humano.
Debe o seu nome a que cando foi descuberto se confundiu cunha bacteria debido ao seu gran tamaño, e a que "imitaba" (en inglés mimic) mediante unha tinguidura de Gram a dunha bacteria grampositiva.
O Mimivirus pertence á nova familia de virus Mimiviridae (mimivíridos), á cal tamén pertence o mamavirus, descuberto posteriormente e aínda de maior tamaño. A familia intégrase dentro dun grupo de virus de orixe monofilética, denominado virus de ADN grande nucleocitoplasmáticos (NCLDV, en inglés) e ao que pertencen virus tan relevantes para a saúde humana e animal como o virus da varíola (familia Poxviridae, poxvíridos), o virus do herpes labial (familia Herpesviridae, herpesvíridos) ou o virus da peste porcina africana ou VPPA (famila Asfarviridae, asfarvíridos).
Realizáronse estudos na Université de le Méditerranée, publicados en Nature, nos que se secuenciou o xenoma do organismo. Os resultados obtidos foron: 800 nm de lonxitude, 1 181 404 pares de bases e 1.262 xenes.
Actualmente estase investigando sobre a posibilidade de que a presenza de mimivirus en pacientes con cadros graves de enfermidade pulmonar estea asociada a un prognóstico negativo; porén, publicáronse datos contraditorios en canto a esta cuestión.[1][2][3] En calquera caso tanto pola súa peculiaridade como pola súa asociación a microorganismos causantes de graves pneumonías infecciosas, trátase dunha entidade modelo de grande interese científico.
Varios membros da comunidade científica declararon recentemente que, debido a que a partícula viral é capaz de xerar as súas propias proteínas, é de feito un organismo vivo, unha idea que non fai más que agudizar a confusión existente con respecto á clasificación dos virus. Mimi, cos seus 911 xenes codificantes, codifica 50 proteínas que nunca antes foran vistas nun virus, como o citocromo P450 implicado no metabolismo enerxético. Ademais codifica proteínas que levan mecanismos bioquímicos que si teñen algúns virus, pero a unha complexidade máxima, como son chaperonas que interveñen no correcto pregamento das proteínas e enzimas que poden proporcionar mecanismos para manter a integridade do ADN (rutas bioquímicas de reparación de erros durante a replicación ou de danos físico-químicos). Representa unha nova familia dentro dos grandes virus ADN nucleocitoplasmáticos que apareceron na Terra hai aproximadamente catro mil millóns de años. Ademais, desafía o sistema de clasificación dos organismos dentro de tres dominios distintos (Archaea, Bacteria e Eukarya), xa que parece exixir a creación dun cuarto dominio. A súa liñaxe é moi antiga e podería ter aparecido antes da existencia dos organismos celulares, o cal pode ter consecuencias importantes nas concepcións sobre a orixe da vida.
O xenoma do mimivirus está conformado por unha molécula de ADN de dupla cadea liñal de 1 181 404 de pares de bases. Isto fixo que fose considerado o maior xenoma viral coñecido, superando ao do miovirus Bacillus phage G (máis tarde descubriuse o Pandoravirus, aínda maior). Ademais do gran tamaño do seu xenoma, estímase que o mimivirus posúe 911 xenes que codifican proteínas, superando considerabelmente os 4 xenes codificantes mínimos requiridos para a existencia dos virus (véxase MSD e virus Qβ). A análise do xenoma do mimivirus mostrou a presenza de xenes non encontrados en ningún ouro virus. Un exemplo é o xene que codifica para a ARNt aminoacil transferase, que até entón se cría exclusivo dos organismos celulares. Como outros virus de gran tamaño, o mimivirus contén varios xenes para o metabolismo de azucres, lípidos e aminoácidos, así como outros xenes relacionados co metabolismo e non encontrados en ningún outro virus. Aproximadamente o 90 % do xenoma é codificante, sendo tan só o 10 % restante "ADN lixo".
O mimivirus posúe un gran número de características que o sitúan na fronteira entre o vivo e o non vivo. É tan grande como algunhas especies de bacterias, tales como Rickettsia conorii e Tropheryma whipplei; o tamaño do seu xenoma é comparábel ao de varias bacterias, incluídas as anteriormente nomeadas, e codifica para produtos que se pensaba que non eran codificados por ningún virus. Ademais, o mimivirus posúe xenes que codifican para a síntesis de nucleótidos e aminoácidos, algúnos dos cales están ausentes incluso en pequenos procariotas intracelulares obrigados. Isto significa que, ao contrario que estas bacterias, o mimivirus non depende do xenoma da célula hospedeira para codificar as rotas metabólicas para estes produtos. Porén, o mimivirus carece de xenes para proteínas ribosómicas, polo que depende da célula hospedeira para a tradución proteica e o metabolismo enerxético.
Todos estes factores, en conxunto, causaron un debate científico sobre se o mimivirus podería ser unha forma de vida distinta, comparábel aos dominios Eukarya, Archaea e Bacteria. Porén, o mimivirus non posúe as seguintes características que forman parte dalgúns dos convenios sobre a definición de vida: homeostase, resposta a estímulos, crecemento no sentido normal do termo (en lugar de replicarse a través do autoensamblamento de compoñentes individuais) ou levar a cabo a división celular.
Debido a que se pensa que podería ter existido con anterioridade aos organismos celulares, o mimivirus ten un papel importante no debate sobre as orixes da vida. Algúns xenes exclusivos do mimivirus, incluíndo aqueles que codifican para a súa cápside, conserváronse nunha gran variedade de virus que infectan organismos de todos os dominios (Eukarya, Archaea e Bacteria). Isto fai pensar que o mimivirus está relacionado cun tipo de virus de ADN que existiu antes que os organismos celulares e que xogou un papel esencial no desenvolvemento da vida na Terra.[4] Unha hipótese alternativa afirma que foron tres tipos distintos de virus de ADN os que deron lugar aos tres dominios coñecidos.[5]
Os partidarios de considerar aos Mimivirus como seres vivos clasifícano como se indica na lista que segue, na que se crea un novo dominio da vida. Os que o clasifican utilizando a clasificación común actual dos virus non introducen os novos taxons superiores (reino, dominio, superdominio).
Os Mimivirus son un grupo de virus de gran tamaño que posúen ADN de cadea dupla e cuxas partículas maduras miden 400 nm de diámetro. O seu ADN posúe 800.000 pares de bases e 900 xenes.
O primeiro foi descuberto en 1992 nunha torre de refrixeración industrial en Bradford (Inglaterra) e identificado en 2003 por un grupo de investigadores da Université de le Méditerranée (Universidade do Mediterráneo) en Marsella (Francia). O virus, que foi descuberto mentres se estudaba a lexionelose (enfermiadade causada poa bacteria Legionella), encontrouse dentro da ameba acuática Acanthamoeba polyphaga. Á súa vez, tamén se encontraron anticorpos para este virus en mostras de sangue humano.
Debe o seu nome a que cando foi descuberto se confundiu cunha bacteria debido ao seu gran tamaño, e a que "imitaba" (en inglés mimic) mediante unha tinguidura de Gram a dunha bacteria grampositiva.
Mimivirus adalah sebuah genus virus yang mengandung satu spesies tunggal bernama Acanthamoeba polyphaga mimivirus (APMV), atau sebuah kelompok virus besar yang saling terkait secara filogenetik (biasanya ditulis MimiN).[1] APMV lebih sering disebut "mimivirus". Sampai Oktober 2011, ketika virus Megavirus chilensis yang lebih besar ditemukan, mimivirus adalah virus yang diameter kapsidnya paling besar.[2] Mimivirus memiliki genom yang besar dan kompleks dibandingkan virus-virus lain. Mimivirus, singkatan dari "mimicking microbe", disebut seperti itu karena ukurannya yang besar dan memiliki sifat pewarnaan Gram yang tampak.[3]
APMV ditemukan secara tidak sengaja pada tahun 1992 di dalam ameba Acanthamoeba polyphaga (dari situlah asal namanya) dalam penelitian Legionellosis. Virus ini diamati melalui pewarna Gram dan salah dikira sebagai bakterium gram-positif. Akibatnya, mikroorganisme ini diberi nama Bradfordcoccus (diambil dari nama distrik asal ameba ini, Bradford, Inggris). Pada tahun 2003, para peneliti Université de la Méditerranée di Marseille, Prancis, menerbitkan hasil penelitian mereka di Science yang mengidentifikasi mikroorganisme ini sebagai virus.[4]
|coauthors= yang tidak diketahui mengabaikan (|author= yang disarankan) (bantuan)
|coauthors= yang tidak diketahui mengabaikan (|author= yang disarankan) (bantuan)
Mimivirus adalah sebuah genus virus yang mengandung satu spesies tunggal bernama Acanthamoeba polyphaga mimivirus (APMV), atau sebuah kelompok virus besar yang saling terkait secara filogenetik (biasanya ditulis MimiN). APMV lebih sering disebut "mimivirus". Sampai Oktober 2011, ketika virus Megavirus chilensis yang lebih besar ditemukan, mimivirus adalah virus yang diameter kapsidnya paling besar. Mimivirus memiliki genom yang besar dan kompleks dibandingkan virus-virus lain. Mimivirus, singkatan dari "mimicking microbe", disebut seperti itu karena ukurannya yang besar dan memiliki sifat pewarnaan Gram yang tampak.
Mimivirus è un genere virale contenente una sola specie ad oggi identificata cui è stato attribuito il nome Acanthamoeba polyphaga mimivirus (APMV), facente parte della famiglia Mimiviridae. Nel linguaggio colloquiale, l'APMV è più comunemente definito con la sola denominazione di “Mimivirus”. Fino alla scoperta del più grande Pandoravirus nel 2013, questa specie possedeva il più grande capside fra tutti i virus conosciuti.[1] Il Mimivirus possiede un genoma più esteso e complesso della maggior parte dei virus.
L'APMV venne scoperto in modo del tutto casuale all'interno di una cellula dell'ameba Acanthamoeba polyphaga, dalla quale ha preso il nome, nel 1992 durante delle ricerche sulla malattia del legionario. Il virus venne identificato attraverso la tecnica della colorazione di gram e, per questo, erroneamente classificato come batterio gram-positivo. In conseguenza di ciò venne chiamato “Bradforcocco” poiché l'ameba dalla quale era stato isolato venne raccolta nel distretto di Bradford, in Inghilterra. Nel 2003, ricercatori della Université de la Méditerranée a Marsiglia, Francia, pubblicarono un articolo sulla rivista “Science” nel quale affermarono di aver identificato il microorganismo come un virus.[2]
Il Mimivirus potrebbe essere l'agente eziologico di alcune forme di polmonite; comunque, questa è una ipotesi basata solamente sull'evidenza della formazione di anticorpi diretti contro questo organismo nel sangue di pazienti colpiti da polmonite.[3] Anche se la classificazione di Mimivirus come un agente patogeno è provvisoria, l'evidenza dimostra una nutrita serie di elementi di prova che può causare la polmonite virale.[4]
Mimivirus è stato inserito dalla Commissione Internazionale per la Tassonomia dei Virus nella famiglia Mimiviridae,[5] e fa parte del gruppo I della classificazione di Baltimore.
Mimivirus appartiene al phylum di grandi virus conosciuto come grandi virus nucleo-citoplasmatici a DNA (NCLDV, nome scientifico Nucleocytoviricota). Essi sono tutti virus di grandi dimensioni che mostrano in comune sia caratteristiche molecolari che genomi molto complessi. Il genoma del Mimivirus possiede inoltre circa 21 geni che codificano per omologhi di proteine che sono state ritrovate con un alto grado di conservazione in quasi tutti i NCLDV, e ricerche successive tenderebbero a mostrare che il Mimivirus sia il membro divergente più recente in questo gruppo.[2]
Il Mimivirus è il virus più grande conosciuto, con un capside del diametro di 400 nanometri. Filamenti proteici della lunghezza di 100 nanometri dipartono dalla superficie del capside, incrementando il diametro totale del virus a 600 nanometri. Molti testi scientifici considerano questa misura molto approssimata, preferendo includere il diametro più probabile dell'organismo in range compreso fra 400 e 800 nanometri dipendentemente da quanto la lunghezza totale del capside e del virione siano misurate. Il capside appare esagonale se visto con un microscopio elettronico, quindi si può ragionevolmente supporre che la simmetria del capside stesso sia icosaedrica. Esso non sembra possedere un pericapside, o envelope, suggerendo che il virus non esca dalla cellula ospite per gemmazione ma che invece ne venga liberato in seguito alla lisi della stessa.[6]
Il mimivirus ha in comune con gli altri NCLDV molte caratteristiche morfologiche. Poiché molti virus di questo gruppo presentano un rivestimento (envelope) lipidico interno circondante il “nucleo” interno, i biologi M. Suzanne-Monti ed altri hanno suggerito che anche questo virus possa presentare una struttura analoga, sebbene non sia ancora stata osservata direttamente. Il denso cuore centrale del virione appare come una regione scura nelle immagini al microscopio elettronico. Il grande genoma del virus risiede completamente in quest'area.
Dalla purificazione dei virioni possono essere isolati molti trascritti di mRNA. Come per altri NCLDV, sono stati riscontrati anche trascritti per la DNA polimerasi, una proteina del capside ed un fattore di trascrizione TFII-simile. Inoltre, tali ricerche hanno mostrato la presenza di tre differenti trascritti di enzimi amminoacil-tRNA sintetasi e quattro molecole sconosciute di RNA tipiche di questo genere di virus. Questi trascritti pre-impacchettati possono essere tradotti in enzimi senza che avvenga una preventiva espressione genica da parte del patrimonio genetico virale e dovrebbero essere necessari per la corretta replicazione del mimivirus. Anche altri virus a DNA, come il citomegalovirus umano e l'Herpesvirus presentano dei trascritti di RNA pre-impacchettati[6].
Un ceppo di Mimivirus conosciuto come Mamavirus si è rivelato il primo virus al mondo a possedere un suo proprio virus parassita, soprannominato "Sputnik"
Il genoma del mimivirus è una molecola lineare e continua di DNA a doppio filamento, contenente 1,2 milioni di coppie di basi. Ciò lo rende il più grande genoma virale mai riscontrato, superando di poco più del doppio il precedente detentore del record, il miovirus “Batteriofago G”. Inoltre, esso è più esteso del genoma di 30 organismi cellulari.[7] In relazione al suo enorme genoma, il mimivirus possiede all'incirca 911 geni codificanti per proteine, superando decisamente il numero minimo di 4 geni che è richiesto ad un virus per poter completare correttamente il ciclo vitale.[8] Analisi del suo genoma hanno mostrato la presenza di geni mai riscontrati in altri virus, inclusi geni codificanti per le amminoacil-tRNA sintetasi, ed altri che erano precedentemente ritenuti come tipici ed esclusivi degli organismi cellulari. Come altri grandi virus a DNA, il mimivirus contiene molti geni coinvolti nel metabolismo di amminoacidi, zuccheri e lipidi, ma anche altri geni metabolici mai riscontrati in altri virus (M. Suzan-Monti, 2006). Circa il 90% del genoma virale ha capacità di codificare, mentre il restante 10% sembrerebbe essere semplice DNA non codificante.
Ciò che fa sì che il mimivirus debba essere considerato un virus e non un batterio, è la modalità di replicazione, non autonoma. Lo svolgimento del processo replicativo del mimivirus non è ancora completamente compreso, ma come minimo è noto che il mimivirus si attacca ad un recettore sulla membrana della cellula dell'ameba e dopo di ciò entra all'interno della stessa per endocitosi. Una volta all'interno, inizia una “fase di eclissi”, durante la quale il virus sparisce e la cellula appare completamente sana e inalterata. Dopo circa quattro ore piccoli accumuli proteici possono essere riscontrati in alcune aree della cellula. Otto ore dopo l'infezione molti virioni di mimivirus risultano chiaramente visibili nella cellula. Il citoplasma della cellula continua a riempirsi di virioni appena sintetizzati e circa 24 ore dopo l'infezione iniziale la cellula probabilmente scoppia rilasciando i nuovi mimivirus.[9]
Poco è conosciuto riguardo ai dettagli della replicazione, probabilmente consistente in attacco alla superficie della cellula ed ingresso, rilascio del genoma conservato nel “core”, trascrizione, traduzione, assemblaggio e rilascio della progenie virale. Tuttavia, gli scienziati hanno stabilito le linee generali del processo utilizzando micrografie elettroniche delle cellule infette in vari momenti dopo l'ingresso del virione. Queste micrografie mostrano che il capside del mimivirus viene assemblato nel nucleo, mentre il DNA acquista un rivestimento interno di membrane lipidiche attraverso gemmazione dalla membrana nucleare, e la presenza di particelle molto simili a quelle prodotte da molti altri virus, compresi tutti gli NCLDV. Queste particelle sono conosciute in altri virus come “fabbriche virali” e contribuiscono ad un efficiente assemblaggio dei virioni attraverso l'alterazione di vaste aree della cellula ospite.
Il mimivirus possiede molte caratteristiche che lo pongono al confine fra viventi e non-viventi. Le sue dimensioni sono analoghe a quelle di molti organismi batterici, come rickettsie e molti altri procarioti, possiede un genoma di dimensioni comparabili a molti procarioti anche non parassiti, che codifica proteine apparentemente inutili per un normale virus In più, esso contiene geni che codificano per enzimi coinvolti nella sintesi di nucleotidi e amminoacidi, che non sono presenti in molti virus parassiti intracellulari obbligati. Ciò significa che differentemente da questi virus il mimivirus non dipende dal genoma della cellula ospite per l'espletazione dei processi metabolici necessari alla loro produzione. Esso tuttavia non possiede i geni per le proteine ribosomiali e dipende perciò dal suo ospite per sintesi proteica e processi energetici. Questi fattori combinati insieme hanno portato gli scienziati a chiedersi se il mimivirus si possa considerare una forma di vita distinta, appartenente ad un ulteriore dominio oltre ai normali eucarioti, batteri ed archea. Nondimeno, il mimivirus non mostra delle caratteristiche che sono considerate essenziali nella corrente definizione di vita: omeostasi, risposta agli stimoli, crescita e riproduzione nel senso classico o divisione cellulare.
Mimivirus è un genere virale contenente una sola specie ad oggi identificata cui è stato attribuito il nome Acanthamoeba polyphaga mimivirus (APMV), facente parte della famiglia Mimiviridae. Nel linguaggio colloquiale, l'APMV è più comunemente definito con la sola denominazione di “Mimivirus”. Fino alla scoperta del più grande Pandoravirus nel 2013, questa specie possedeva il più grande capside fra tutti i virus conosciuti. Il Mimivirus possiede un genoma più esteso e complesso della maggior parte dei virus.
Mimivirus is een geslacht van virussen. Het bevat maar één soort: Acanthamoeba polyphaga mimivirus (APMV).
Het is het grootste virus tot zover bekend, groter dan bepaalde bacteriën. De capside van het virus heeft een diameter van 400 nm. Het virus werd in 1992 ontdekt in de amoebe Acanthamoeba polyphaga bij een onderzoek naar legionairsziekte in een koeltoren in Bradford. Eerst werd het aangezien voor een gram-positieve bacterie wegens zijn grootte en zijn gelijkaardigheid aan bepaalde bacteriën. Het kreeg de naam Bradfordcoccus. Toen men zich de fout had gerealiseerd werd de ontdekking, die dus een virus bleek te zijn, hernoemd tot Mimicking microbe virus of kortweg Mimivirus.
Het virus onderscheidt zich van andere virussen door zijn grote genoom. Daarnaast codeert het virus voor bepaalde producten die niet eerder bij een ander virus zijn waargenomen. Zo bezit het mimivirus genen die coderen voor nucleotide- en aminozuursynthese. Dit zijn genen die zelfs bepaalde intracellulaire bacteriën niet hebben. Hierdoor is ook de kwestie van non-cellulair leven meer ter sprake gebracht.
Mimivirus behoort tot de dsDNA-virussen en wordt bij de groep van de nucleocytoplasmische grote DNA-virussen gerekend, waar ook de families Poxviridae, Iridoviridae, Phycodnaviridae, Asfarviridae en Coccolithoviridae toebehoren.
Mimivirus is een geslacht van virussen. Het bevat maar één soort: Acanthamoeba polyphaga mimivirus (APMV).
Het is het grootste virus tot zover bekend, groter dan bepaalde bacteriën. De capside van het virus heeft een diameter van 400 nm. Het virus werd in 1992 ontdekt in de amoebe Acanthamoeba polyphaga bij een onderzoek naar legionairsziekte in een koeltoren in Bradford. Eerst werd het aangezien voor een gram-positieve bacterie wegens zijn grootte en zijn gelijkaardigheid aan bepaalde bacteriën. Het kreeg de naam Bradfordcoccus. Toen men zich de fout had gerealiseerd werd de ontdekking, die dus een virus bleek te zijn, hernoemd tot Mimicking microbe virus of kortweg Mimivirus.
Het virus onderscheidt zich van andere virussen door zijn grote genoom. Daarnaast codeert het virus voor bepaalde producten die niet eerder bij een ander virus zijn waargenomen. Zo bezit het mimivirus genen die coderen voor nucleotide- en aminozuursynthese. Dit zijn genen die zelfs bepaalde intracellulaire bacteriën niet hebben. Hierdoor is ook de kwestie van non-cellulair leven meer ter sprake gebracht.
Mimivirus behoort tot de dsDNA-virussen en wordt bij de groep van de nucleocytoplasmische grote DNA-virussen gerekend, waar ook de families Poxviridae, Iridoviridae, Phycodnaviridae, Asfarviridae en Coccolithoviridae toebehoren.
Mimivirus – największy znany dwuniciowy wirus DNA, którego kapsyd, będący dwudziestościanem foremnym, ma średnicę dochodzącą do 400 nm. Zsekwencjonowanie w 2004 roku ujawniło, że posiada wyjątkowo duży jak na wirusa genom (1 181 404 par zasad) z 1258 genami[1].
Został odkryty w 1992 roku w przemysłowej wieży chłodniczej w Bradford, a zidentyfikowany w 2003 przez badaczy z Université de la Méditerranée w Marsylii. Sam wirus został znaleziony we wnętrzu Acanthamoeba polyphaga podczas badań nad legionellozą.
Mimivirus – największy znany dwuniciowy wirus DNA, którego kapsyd, będący dwudziestościanem foremnym, ma średnicę dochodzącą do 400 nm. Zsekwencjonowanie w 2004 roku ujawniło, że posiada wyjątkowo duży jak na wirusa genom (1 181 404 par zasad) z 1258 genami.
Został odkryty w 1992 roku w przemysłowej wieży chłodniczej w Bradford, a zidentyfikowany w 2003 przez badaczy z Université de la Méditerranée w Marsylii. Sam wirus został znaleziony we wnętrzu Acanthamoeba polyphaga podczas badań nad legionellozą.
Mimivirus é uma espécie de vírus explosivo, parasita obrigatório do momozoário (Classe Rhyzopoda ou Sarcodina) Acanthamoeba polyphaga, por isso seu nome científico é Acanthamoeba polyphaga mimivirus (APMV). Popularmente o APMV é mais conhecido apenas como “mimivírus”.
Foi descoberto em 1992 na Acanthamoeba polyphaga, porém, na época, acreditava-se ser uma bactéria do gênero Legionella. O vírus foi observado através de coloração do tipo Gram, por isso foi erradamente considerado uma bactéria gram-positiva. A consequência deste erro foi o nome que lhe deram, "Bradfordcoccus", devido ao distrito onde foi recolhido a ameba (Bradford na Inglaterra). Em 2003, pesquisas da Université de la Méditerranée em Marseille, França, publicaram uma página na revista Science identificando este microrganismo como sendo um vírus.[1]
O mimivírus já foi considerado um possível causador de algumas formas de pneumonia, já que foram encontrados anticorpos para este vírus em alguns indivíduos com pneumonia.[2] Atualmente acredita-se que ele infecta apenas as amebas, e estas causam a pneumonia.[carece de fontes?]
O mimivírus tem um capsídeo protéico no tamanho de 390 nm (nanômetros) e os filamentos protéicos de 100 nm, totalizando 600 nm, é um dos maiores vírus conhecidos (ficando atrás dos mamavirus e do Megavirus chilensis), sendo maior que algumas bactérias (que possuem 400 nm).
Através de microscopia eletrônica, nota-se que ele tem forma hexagonal, portanto seu capsídeo tem simetria icosaédrica,[3] o que sugere que o vírus não sai da célula parasitada através de exocitose.[4]
O sequenciamento do seu genoma foi feita pelo grupo de Jean-Michel Claverie e, através deste estudo, características inéditas foram vistas neste vírus.
O mimivírus contém tanto o DNA quanto RNA, ao contrário dos vírus "normais". O tamanho do genoma é de 1 181 404 pares de bases,[5] maior do que o de muitas bactérias (algumas bactérias têm apenas 400 genes), ele contém 911 genes codificadores de proteínas, muito acima do número mínimo de 4 genes necessários para um vírus existir.[6] Análises do seu genoma identificaram genes nunca antes vistos em outros vírus, inclusive genes responsáveis pelo metabolismo de açúcares, lípidos e aminoácidos, inclusive genes responsáveis pela produção de sintetizadores de RNAt.[4]
O mimivírus tem a capacidade de produzir proteínas pelos seus próprios meios e a capacidade de reparação do seu próprio genoma, sendo que todas estas propriedades nunca foram vistas em outros vírus. No mimivirus 90% de seus genes são utilizáveis, apenas 10% é Homeobox (Arquivo Genético, Passado Génico, ou "Entulho Genético"), ao contrário da maioria dos organismos.
O gigantesco Mimivírus se defende de invasores utilizando semelhante ao sistema CRISPR implementado por bactérias e outros microorganismos.[7] O sistema de defesa do Mimivírus pode levar a descoberta de novas ferramentas de edição de genoma[8].
Mimivirus é uma espécie de vírus explosivo, parasita obrigatório do momozoário (Classe Rhyzopoda ou Sarcodina) Acanthamoeba polyphaga, por isso seu nome científico é Acanthamoeba polyphaga mimivirus (APMV). Popularmente o APMV é mais conhecido apenas como “mimivírus”.
APMV був випадково знайдений в 1992 році в амебі Acanthamoeba polyphaga, на честь якої він і був названий, під час дослідження легіонельозу. Вірус був виявлений в препараті забарвленням за Грамом, і внаслідок цього був помилково прийнятий за грампозитивну бактерію. Цей організм був названий Bradfordcoccus на честь району в якому була знайдена амеба (Бредфорд, Англія). У 2003 році дослідники з Середземноморського університету (Марсель, Франція) опублікували статтю в журналі Science, у якій описали даний мікроорганізм як вірус[1].
Існує гіпотеза, що мімівірус може викликати у людей деякі форми пневмонії. Досі були знайдені лише непрямі свідоцтва на користь цієї гіпотези у вигляді антитіл до вірусу, виявлених у пацієнтів, що страждають на цю хворобу[2][3].
Мімівірус не був досі поміщений Міжнародним комітетом з таксономії вірусів до будь-якої родини, але, на підставі даних отриманих при дослідженні метагеному, передбачається існування додаткових членів запропонованої родини Mimiviridae[4]. Згідно з класифікацією за Балтимором, мімівірус був віднесений до групи I, тобто групи вірусів, чий генетичний матеріал являє собою дволанцюгову ДНК, і що не мають РНК-стадії. До цієї групи входять такі родини вірусів як ірідовіруси, поксвіруси та інші. Всі ці віруси відрізняються великими розмірами, схожими молекулярними характеристиками і складними геномами[1].
Ряд білків мімівірусу, що беруть участь в реплікації генома, виявились гомологічними білкам інших великих нуклеоплазматичних ДНК-містячих вірусів (поксвіруси, ірідовіруси, фікоднавіруси), що говорить про їх загальне походження. Проте велика кількість мімівірусних білків не виявляє схожість ні з одним відомим в наш час[коли?] білком. Крім того, геном мімівіруса кодує значну кількість білків, що нагадують еукаріотичні і бактеріальні. Ймовірно, як це часто трапляється з вірусами, ці гени були придбані мімівірусом повторно і походять з геномів хазяїв вірусу та їх паразитів[5].
Мімівірус, маючи капсид діаметром 400 нм з численними 100-нанометровими білковими нитками на ньому, є одним із найбільших вірусів серед відомих на сьогоднішній день (найбільший відомий вірус це Pithovirus). У науковій літературі приведені розміри віріону від 400 нм до 800 нм, залежно від того, чи заміряється діаметр капсида або загальна довжина вірусу в подовжній осі. Капсид містить три шари щільної речовини, ймовірно двох ліпідних мембран товщиною 4 нм кожна та білкової оболонки, товщиною близько 7 нм[6]. Під електронним мікроскопом можна спостерігати гексагональну форму капсида, що указує на ікосаедральну симетрію білкових структур капсида. У мімівірусу не спостерігається зовнішньої оболонки, що указує на те, що мімівірус покидає заражену клітку не шляхом екзоцитозу[7].
Мімівірус володіє багатьма особливостями будови, характерними і для інших вірусів своєї групи. Ліпідний шар, яким вистилається внутрішня поверхня капсиду і який був знайдений у всіх цих вірусів, за припущенням М. Сьюзан-Монті (M. Suzan-Monti) та співавторів[8] також присутній і у мімівірусу. Центральна частина вірусу, що містить в собі ДНК, виглядає під електронним мікроскопом як темнозабарвлена область.
З очищених віріонів були виділені декілька різних мРНК, що кодують ДНК-полімеразу, білки капсиду і фактори транскрипції, близькі до TFII. Також були знайдені мРНК, що кодують аміноацил-тРНК-синтетазу і 4 неідентифікованих молекули мРНК, специфічних для мімівірусу. Ці мРНК можуть бути трансльовані без експресії вірусних генів і, ймовірно, необхідні мімівірусу для реплікації. Інші ДНК-вмісні віруси, такі як і вірус герпесу (Herpes simplex virus type-1), також містять подібні мРНК[8].
Геном мімівірусу складається з лінійної молекули ДНК, що містить близько 1 185 000 пар основ[9]. Це найбільший геном серед всіх відомих науці вірусів, він в два рази довший, ніж наступний за розміром геном міовірусу Bacillus phage G. Крім того, мімівірус володіє найбільшим об'ємом генетичної інформації, ніж як мінімум 30 організмів, що мають клітинну будову[6].
Додатково до унікального для вірусів розміру геному, мімівірус володіє приблизно 911 генами, що кодують білки, що значно більше 4 необхідних кожному вірусу генів[10]. Аналіз геному показав наявність генів, не присутніх ні у яких інших вірусів, зокрема генів, що кодують аміноацил-тРНК-синтетазу та інших, виявлених тільки у організмів з клітинною будовою. Як і інші великі ДНК-вмісні віруси, мімівірус містить набір генів для кодування ферментів вуглеводного, ліпідного і амінокислотного метаболізму, проте серед них є і такі, які не знайдені у інших вірусів[8].
Приблизно 10 % геному припадає на некодуючі ділянки ДНК.
Гіпотетично представники групи великих нуклеоплазматичних ДНК-вмісних беруть свій початок від складніших (можливо клітинних) форм, таких як сучасні мікоплазми і рікетсії, тобто бактерій, що в процесі еволюції в результаті паразитичного способу життя втратили велику частину генів. На користь цієї гіпотези говорить наявність в геномі цих вірусів великої кількості «надмірних» генів, що не є необхідними для розмноження, і функціонально дублюють гени хазяїв. Вважається, що ці гени є свого роду «реліктом», що залишився від періоду «самостійності». Багато з вірусних генів не має гомологів в геномах клітинних організмів, що указує на еволюційну відокремленість їх гіпотетичного предка від архей, бактерій і еукаріотів, що теоретично дозволяє виділити їх до четвертого домену життя.
Хоча ряд білків мімівірусу, що беруть участь в реплікації генома, і є гомологічними білкам деяких інших вірусів, мімівірус виявився свого роду рекордсменом за кількістю унікальних генів, що не мають схожості ні з одним білком, відомим в даний час, і тому деякі учені вважають мімівірус генетичним реліктом, близьким до загального предка великих ДНК-містячих вірусів. З вивченням мімівірусу пов'язується надія отримати інформацію про цього загального предка.
Ще однією цікавою властивістю генома мімівірусу виявилася наявність великої кількості гомологічних копій одних і тих же генів. Мабуть, деякі гени предка мімівіруса піддалися дуплікації, а потім еволюціонували незалежно один від одного. Це спостереження дозволило деяким ученим припустити, що екстраординарні розміри генома мімівірусу пояснюються не стільки його близькістю до гіпотетичного предка, скільки особливостями займаної ним екологічної ніші, що накладає менші обмеження на розміри геному[5].
Стадії реплікації мімівіруса все ще слабо вивчені. Відомо, що мімівірус приєднується до рецепторів на поверхні клітин амеби і за допомогою них потрапляє всередину клітини. Всередині вірус розпадається, а інфікована клітина продовжує нормальну життєдіяльність. Приблизно через 4 години в декількох місцях усередині амеби починають з'являтися скупчення не характерної для неї речовини, а через 8 годин після інфекції в клітці вже добре помітна безліч віріонів мімівіруса. Цитоплазма продовжує наповнюватися новими віріонами і через 24 години після інфекції клітина розривається і вивільняє вірусні частинки[11].
Дуже небагато відомо про деталі життєдіяльності вірусу всередині клітини, зокрема процесу реплікації, транскрипції, трансляції та збірки віріонів. Проте за допомогою електронної мікроскопії інфікованих клітин була виявлена агрегація капсидів мімівіруса у ядрі клітини та встановлено утворення внутрішнього ліпідного шару вірусу відбрунькуванням від ядра, а також знайдені частинки, схожі з «вірусними фабриками», які утворюються при інфекціях іншими вірусами тієї ж групи.
Мімівірус володіє багатьма властивостями, які поміщають його на межу живого і неживого. За своїми розмірами він перевершує деяких бактерій, таких як Rickettsia conorii або Tropheryma whipplei, містить геном, що має розміри, порівняні з розмірами геномів багатьох паразитичних бактерій (зокрема вищеназваних), і має гени, не знайдені у інших вірусів, зокрема що такі, кодують ферменти синтезу нуклеотидів і амінокислот, які відсутні навіть у деяких дрібних бактерій-внутріклітинних паразитів. Це означає відносну незалежність мімівіруса (на відміну від вказаних бактерій) від геному клітини-хазяїна, що кодує основні метаболічні шляхи. Проте мімівірус не має генів синтезу рибосомних білків, через що він має необхідність в рибосомах хазяїна. Поєднання цих властивостей викликало в науковому середовищі суперечки, чи є мімівірус особливою формою життя, доменом, разом з еукаріотами, бактеріями і археями[12].
Проте, мімівірус не володіє гомеостазом, не відповідає на подразники, не росте і не розмножується самостійно (натомість синтезується клітиною і самозбирається в ній з окремих компонентів), що типово для вірусів.
Гени, властиві мімівірусу (зокрема білки, що кодують капсид), зберігаються в безлічі вірусів, що вражають організми всіх трьох доменів. На підставі цього факту робиться припущення, що мімівірус пов'язаний з ДНК-містячими вірусами, які з'явилися приблизно одночасно з найстародавнішими організмами, що мають клітинну будову, і займають ключове положення в походженні життя на Землі[13].
|issie= (довідка) |issie= (довідка) Mimivirus là một chi virus bao gồm một loài được khám phá ra, loài này có tên khoa học Acanthamoeba polyphaga mimivirus (APMV). APMV thường được gọi một cách đơn giản là "mimivirus", có nghĩa "virus bắt chước". Nó có vỏ capsid với đường kính lớn nhất của các virus được khám phá, cũng như bộ gen lớn và phức tạp đối với những virus khác. Tuy những nhà sinh học chưa hiểu biết nhiều về virus này, nhưng cộng đồng sinh học chú trọng khám phá này vì cấu trúc phức tạp của nó, có cả người miêu tả nó là vực mới và liên kết chờ đợi dã lâu giữa virus và vi khuẩn.
APMV tình cờ được khám phá ra năm 1992 ở trong amip Acanthamoeba polyphaga, nguồn tên của nó, trong việc nghiên cứu về bệnh legionellosis. Virus này được tìm thấy sau khi nhuộm Gram, nên nhà khoa học tưởng lầm nó là vi khuẩn Gram dương. Bởi vậy, nó được đặt tên là "Bradfordcoccus", theo nguồn của amip tại Bradford (Anh). Năm 2003, những nhà nghiên cứu tại trường Đại học Địa Trung Hải tại Marseille (Pháp) xuất bản bài thuyết trình trong tạp chí Science nhận ra vi sinh vật này là loài virus.[1]
Mimivirus có thể là tác nhân gây ra một số loại viêm phổi; tuy nhiên, đây là đề xuất lâm thời chỉ dựa trên bằng chứng gián tiếp theo những kháng thể gặp trong người bị viêm phổi.[2] Tuy mimivirus chỉ được phân loại là tác nhân gây bệnh tạm thời, nhưng càng ngày càng thêm bằng chứng rằng nó có thể gây viêm phổi virus.[3]
Ủy ban Quốc tế về Phân loại Virus (International Committee on Taxonomy of Viruses, ICTV) chưa xếp mimivirus trong họ virus. Tuy nhiên, người ta dự đoán theo dữ liệu siêu bộ gen (metagenomic) rằng họ được đề xuất, Mimiviridae, bao gồm thêm loài.[4] Ngoài ra, nó được xếp trong Nhóm I của hệ thống phân loại Baltimore.
Mimivirus thuộc về một nhóm virus cỡ lớn, virus DNA lớn có nhân và tế bào chất (nucleocytoplasmic large DNA viruses, NCLDV), đây không phải là hệ thống phân loại theo đúng nghĩa. Các virus này là loài lớn đều có đặc tính phân tử chung và bộ gen lớn. Bộ gen mimivirus cũng chứa 21 gene để biến chất đồng đãng thành protein mà phần nhiều NCLDV vẫn giữ rất nhiều, và sự nghiên cứu gợi ý rằng mimivirus là một loài tách ra sớm khỏi nhóm chung NCLDV.[1]
Mimivirus là một trong những virus lớn nhất được khám phá, chỉ có loại phụ mamavirus thì lớn hơn, với đường kính vỏ capsid tới 400 nm. Những sợi protein dài đến 100 nm nhô ra từ mặt capsid, tính vào những sợi này thì virus dài đến 600 nm tổng cộng. Những con số này rất sơ sơ; các thuyết trình khoa học nói khác nhau, trong khi "cỡ" của virus được cho là từ 400 nm đến 800 nm, tùy theo tính chiều dài tổng cộng hay đường kính capsid. Với chiều dài này, nó lớn bằng vài loài vi khuẩn, như là Rickettsia conorii (bệnh còi xương) and Tropheryma whipplei (bệnh Whipple).[5] Capsid hình như có lục giác khi nhìn dưới kiếnh hiển vi điện tử, cho nên tính đối xứng capsid có hình khối hai mươi mặt đều. Hình như nó không có vỏ virus bên ngoài, nên có lẽ virus không thoát khỏi tế bào chủ bằng phương pháp xuất bào.[6]
|coauthors= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp) |coauthors= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp) |coauthors= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp) |month= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Kiểm tra giá trị ngày tháng trong: |date= (trợ giúp) |coauthors= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp) |coauthors= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp)
|coauthors= bị phản đối (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp); Đã định rõ hơn một tham số trong |author= và |last= (trợ giúp)
Mimivirus là một chi virus bao gồm một loài được khám phá ra, loài này có tên khoa học Acanthamoeba polyphaga mimivirus (APMV). APMV thường được gọi một cách đơn giản là "mimivirus", có nghĩa "virus bắt chước". Nó có vỏ capsid với đường kính lớn nhất của các virus được khám phá, cũng như bộ gen lớn và phức tạp đối với những virus khác. Tuy những nhà sinh học chưa hiểu biết nhiều về virus này, nhưng cộng đồng sinh học chú trọng khám phá này vì cấu trúc phức tạp của nó, có cả người miêu tả nó là vực mới và liên kết chờ đợi dã lâu giữa virus và vi khuẩn.
Мимивирус имеет примерно икосаэдрический капсид диаметром 400—500 нм[13][19]. Капсид покрыт многочисленными белковыми нитями длиной 80—120 нм. В научной литературе приводятся размеры вириона от 400 до 800 нм, в зависимости от того, замеряется ли диаметр капсида или общая длина вируса вдоль продольной оси, включая белковые нити. Основной белок капсида мимивируса является продуктом гена L425 и состоит из двух доменов с укладкой типа jelly-roll («рулет»). Этот белок формирует гомотримерные капсомеры — единицы организации капсида. Капсомеры упакованы гексагонально в виде «ромашек»: шесть капсомеров окружают одно углубление между ними[18]. Также в составе капсида обнаруживается структурный белок сердцевины L410[9].
На одной из вершин капсида обнаружена звёздчатая структура, лучи которой разделяют пять треугольных граней, сходящихся в этой вершине. Лучи имеют ширину около 50 нм, толщину — 40 нм и длину — 200 нм, почти доходя до соседних вершин. Присутствие этой структуры изменяет взаимное расположение граней вириона, в результате чего его форма отклоняется от идеально икосаэдрической: через вирион можно провести только одну ось пятилучевой симметрии, которая проходит через вершину, отмеченную звёздчатой структурой. Так как на поверхности звёздчатой структуры не наблюдается гексагонально упорядоченных углублений, предполагают, что она состоит из белка, который отличается от основного белка капсида[18]. Данная структура играет особую роль при заражении клетки-хозяина: при инфицировании звёздчатая «застёжка» открывается, и через это место ДНК вируса выходит из капсида. По этой причине звёздчатую структуру ещё называют «звёздными вратами»[20].
У мимивируса не наблюдается внешней оболочки, что указывает на то, что он покидает заражённую клетку не путём экзоцитоза[21].
Капсид мимивируса покрыт снаружи густым слоем длинных белковых нитей. Исследование этих нитей с помощью атомно-силового микроскопа показало, что они часто бывают присоединены к общей несущей структуре и заканчиваются небольшой глобулой. Однако до сих пор не известно, к каким участкам поверхности капсида они прикрепляются[18]. Белковые нити устойчивы к воздействию протеаз до тех пор, пока их не обработать лизоцимом, это указывает на то, что эти нити покрыты пептидогликаном. Это хорошо согласуется с тем, что мимивирус окрашивается по методу Грама. Существует мнение, что сильно гликозилированная поверхность нитей может играть роль в привлечении амёб-хозяев[19].
Мимивирус обладает многими особенностями строения, характерными для других крупных ядерно-цитоплазматических ДНК-содержащих вирусов. Например, сразу под капсидом у мимивируса находятся два электронноплотных слоя, которые предположительно являются мембранами[19]. Под этими мембранами находится белковая оболочка толщиной около 7 нм, внутри которой заключена линейная двухцепочечная ДНК вируса. Все описанные компоненты образуют так называемый нуклеокапсид. Стенки нуклеокапсида отстают от стенок капсида примерно на 30 нм, в районе звёздчатой структуры поверхность нуклеокапсида вдавлена[18]. Предполагают, что пространство между вершиной звёздчатой структуры и нуклеокапсидом может быть заполнено гидролитическими ферментами, которые нужны для проникновения вируса в клетку. Между капсидом и нуклеокапсидом были обнаружены внутренние белковые нити, которые предположительно обеспечивают стабильное положение второго внутри первого[19].
Помимо структурных белков капсида, в составе вириона обнаруживают и другие белки, которые классифицируют на несколько функциональных групп:
Кроме белков и ДНК, из очищенных вирионов были выделены несколько различных мРНК, кодирующих ДНК-полимеразу (R322), главный белок капсида L425, TFII-подобный фактор транскрипции (R339), 3 аминоацил-тРНК-синтетазы (L124, L1164 и R663) и 4 неидентифицированных белка, специфичных для мимивируса. По-видимому, трансляция этих мРНК клеточным аппаратом биосинтеза белка необходима для начала репликации вируса. Другие ДНК-содержащие вирусы, такие как цитомегаловирус (Cytomegalovirus) и вирус простого герпеса (Herpes simplex virus type-1), также содержат мРНК[21].
Геном мимивируса, состоящий из линейной молекулы двухцепочечной ДНК, был полностью секвенирован в 2004 году[22][23]. Он содержит 1 181 404 пары оснований и является вторым крупнейшим известным геномом среди вирусов, уступая только Megavirus chilensis (по данным на 2012 год)[24]. Кроме того, мимивирус обладает бо́льшим объёмом генетической информации, чем как минимум 30 организмов, имеющих клеточное строение[25].
По сведениям на 2010 год в геноме мимивируса насчитывают 986 открытых рамок считывания, 6 из которых кодируют тРНК[22][26][27]. Такое количество белковых продуктов является очень большим для вируса — некоторые вирусы обходятся всего четырьмя минимально необходимыми белками[28]. Детальные исследования генома всё ещё продолжаются: исправляются ошибки секвенирования, обнаруживаются новые рамки считывания[29].
Несмотря на большой размер генома, он используется довольно эффективно: на долю кодирующей ДНК приходится около 90,5 % всего объёма генома, это близко к тому, что наблюдается у других крупных ядерно-цитоплазматических ДНК-содержащих вирусов. Открытые рамки считывания разделены между собой промежутками примерно в 157 пар нуклеотидов. Две цепочки ДНК, названные R (англ. right — правая) и L (англ. left — левая), кодируют примерно одинаковое количество генов (450 и 465 соответственно, по данным на 2010 год). В геноме мимивируса не обнаруживается признаков деградации, характерных для геномов паразитических бактерий, таких как псевдогены и мобильные генетические элементы. Содержание адениновых и тимидиновых нуклеотидов довольно высоко — 72 %, что ведёт к повышенному содержанию в белках аминокислот, кодируемых АТ-богатыми кодонами (например, изолейцина, аспарагина и тирозина). Вблизи концов молекулы ДНК были обнаружены инвертированные повторы длиной 617 пар нуклеотидов. Предполагают, что комплементарное взаимодействие этих участков может приводить к формированию Q-структуры — кольцевой ДНК с двумя небольшими хвостами[29].
Примерно для половины генов мимивируса не обнаружены гомологи в современных базах данных, и только для 24 % известна предполагаемая функция[27].
В геноме мимивируса были обнаружены гомологи практически всех ключевых генов, характерных для других крупных ядерно-цитоплазматических вирусов. В то же время данный геном содержит примерно вдвое больше генов, чем геномы родственных вирусов, причём среди этих генов много уникальных. Например, геном мимивируса кодирует несколько белковых компонентов аппарата трансляции: тирозил-, аргинил-, цистеил- и метионил-тРНК-синтетазы, гомологи факторов инициации трансляции eIF4E (L496), eIF4A (R458) и SUI1/eIF1 (R464), фактора элонгации трансляции eEF-1 (R624) и фактора терминации трансляции eRF1 (R726). Помимо генов белков, участвующих в трансляции, обнаружено 6 генов, которые предположительно кодируют тРНК, узнающие кодоны для лейцина, триптофана, гистидина и цистеина. Кроме того, мимивирус кодирует два гомолога РНК-урацил-5-метилтрансферазы (R405, R407) — фермента, метилирующего остаток урацила в тРНК и рРНК[29].
К другим необычным для вирусов генам относятся гены трёх типов топоизомераз и полного набора ферментов репарации, которые способны исправлять ошибки в ДНК, возникшие в результате действия окислителей, ультрафиолетового облучения и алкилирующих веществ. Также мимивирус кодирует ферменты углеводного, липидного и аминокислотного метаболизма[9][30].
Механизмы приспособления мимивируса к изменениям среды на протяжении многих поколений демонстрируют, наряду с признаками дарвинистической эволюции, признаки, соответствующие принципам ламаркизма. Так, например, у мимивируса в условиях пониженной конкуренции происходит репрессия некоторых генов. Это изменение наследуется следующими поколениями вируса, в некоторых случаях усиливаясь вплоть до полной инактивации этих генов. Предположительно, эффект является следствием худшей репарации менее используемых генов[31].
Мимивирус является одним из немногих дцДНК-вирусов, в геноме которых обнаружена последовательность, кодирующая интеин. Интеин представляет собой белковый домен, который катализирует своё собственное вырезание из несущей молекулы и сшивание образовавшихся концов. Такая последовательность присутствует в гене ДНК-полимеразы B мимивируса[32].
Октамерная последовательность AAAATTGA была обнаружена примерно перед половиной генов мимивируса в положении от −80 до −50. Эта последовательность является TATA-подобным промоторным элементом и узнаётся транскрипционным аппаратом вируса на ранней стадии жизненного цикла[9]. Другая вырожденная АТ-богатая последовательность является поздним промотором[26].
Первым известным хозяином мимивируса является амёба Acanthamoeba polyphaga. Попытки экспериментально инфицировать клетки других одно- и многоклеточных организмов показали, что только другие представители рода Acanthamoeba — A. castellanii и A. mauritaniensis — могут служить хозяевами данного вируса[21]. Некоторые данные указывают на то, что мимивирус может проникать в макрофаги и размножаться в них[6].
Мимивирус имеет 24-часовой литический (сопровождающийся лизисом клетки-хозяина) жизненный цикл с эклипс-фазой продолжительностью 4—5 часов[21]. Все стадии жизненного цикла протекают в цитоплазме клетки[34].
Заражение амёбы мимивирусом предположительно происходит по следующему сценарию:
Существует гипотеза, что мимивирус может вызывать у людей некоторые формы пневмонии[6]. До сих пор были найдены лишь косвенные свидетельства в пользу этой гипотезы. Во-первых, было показано, что в условиях эксперимента мимивирус способен заражать человеческие макрофаги, проникая в клетки в процессе фагоцитоза, и реплицироваться в них[1]. Во-вторых, в нескольких исследованиях у небольшого количества пациентов, страдающих пневмонией, были обнаружены антитела против мимивируса[36][37]. Также был описан единичный случай развития пневмонии у лаборанта, который работал с культурами этого вируса. Содержание антител против мимивируса в его крови тоже было повышенным[38]. Тем не менее, присутствие антител против вируса само по себе не говорит о его патогенности, не исключено, что мимивирус просто обладает сильными иммуногенными свойствами[29].
С другой стороны, ни в одном из зарегистрированных случаев не удалось выделить вирус в чистом виде из образцов жидкостей, полученных от пациентов. Кроме того, исследования, в которых использовалась полимеразная цепная реакция, не обнаружили присутствия мимивируса у пациентов с пневмонией. В 2012 году группа Ванспоуна опубликовала результаты исследования больных пневмонией с целью определить роль мимивируса как возможного патогена. Ни у одного из 109 исследованных пациентов не был обнаружен мимивирус, и лишь у трёх были обнаружены антитела к нему[39]. В целом, вопрос о патогенности мимивируса для человека остаётся открытым, но в качестве охранной меры предлагается рассматривать его как организм II группы патогенности[29].
Научная группа, открывшая мимивирус, также изолировала несколько других родственных ему вирусов, включая чуть более крупный мамавирус[en] (англ. Mamavirus). При изучении вирусных фабрик мамавируса было обнаружено, что на них также собираются небольшие вирионы другого вируса, который был назван спутником (англ. Sputnik)[40]. Спутник, по всей видимости, сам не способен заражать клетки амёб и размножаться в них, но может делать это совместно с мама- или мимивирусом, что классифицирует его как вирус-сателлит. Спутник стал первым известным вирусом-сателлитом, содержащим двухцепочечную ДНК и размножающимся в эукариотических клетках. Однако авторы работы предлагают рассматривать спутник не просто как сателлит, а как вирофаг (вирус вируса) по аналогии с бактериофагами (вирусами бактерий)[41][42][27]. Разница в этих двух концепциях заключается в том, что вирусы-сателлиты полагаются на другой вирус и клетку-хозяина для своего размножения. Вирофаги же, как предполагается, воспроизводятся только за счёт репликативного аппарата вируса-хозяина, то есть являются паразитами исключительно другого вируса[29]. Хотя строгое доказательство ещё предстоит получить, некоторые факты говорят в пользу того, что спутник действительно является вирофагом. Например, в его геноме присутствуют регуляторные элементы, характерные для мимивируса и узнаваемые его транскрипционным аппаратом (последовательности, близкие к позднему промотору мимивируса, сигналы полиаденилирования). Кроме того, присутствие спутника снижает продуктивность размножения мимивируса: лизис клетки-хозяина происходит позже, образуются дефектные вирионы[40].
На сегодняшний день был открыт второй вирофаг мимивируса штамма CL[43].
Мимивирус и другие крупные ядерно-цитоплазматические ДНК-содержащие вирусы обладают рядом свойств, которые не вписываются в традиционные представления о вирусах: крупный размер вирионов, присутствие в вирионе одновременно двух типов нуклеиновых кислот, большой размер и сложность генома, присутствие нехарактерных для вирусов генов (гены белков, участвующих в трансляции, репарации ДНК и фолдинге белков) и возможность выступать в качестве хозяина для другого вируса[14]. Эти факты возродили интерес к вопросу о происхождении и эволюции вирусов.
Было высказано две принципиально разных гипотезы о происхождении сложного генома мимивируса. Согласно первой из них, мимивирус и другие крупные ядерно-цитоплазматические ДНК-содержащие вирусы произошли от более сложного предка (клетки или вируса) путём редукционной эволюции и принадлежат к четвёртому домену живого[22]. По второй гипотезе, эволюция этих вирусов шла по пути усложнения и постепенного накопления генов, поступающих из геномов других организмов путём горизонтального переноса[44]. Фактически обе гипотезы опираются на данные сравнительной геномики и протеомики, которые могут быть интерпретированы по-разному, учитывая тот факт, что они отражают события, возможно, происходившие сотни миллионов лет назад. Относительно быстрые темпы эволюции вирусов и интенсивный горизонтальный перенос генов осложняет анализ и затрудняет построение филогенетических деревьев. Собственно факты, с которыми согласно большинство исследователей, заключаются в том, что более половины генов (и мотивов укладки белков) мимивируса не имеют известных гомологов. Другие данные и их интерпретация продолжают оставаться темой горячих споров.
В геноме мимивируса были обнаружены необычные для вирусов гены, ортологи которых присутствуют у организмов всех трёх доменов (гены аминоацил-тРНК-синтетаз, субъединиц РНК- и ДНК-полимераз). Анализ этих и других генов мимивируса, для которых известны гомологи, позволил определить его относительное положение на филогенетическом дереве. Однако, в зависимости от алгоритма анализа, были получены очень разные результаты. По мнению одних исследователей, линия, ведущая к современному мимивирусу, обособилась примерно в то же время, что и эукариотическая, или даже раньше[22][24]. Сходные результаты были получены и при сравнительном анализе типов белковых укладок[45]. Другие же учёные настаивают на том, что эти гены были приобретены мимивирусом в результате горизонтального переноса от представителей эу- и прокариот и что нет никаких оснований выделять вирусы в четвёртый домен живого[46][47]. Кроме того, большой размер генома мимивируса может быть объяснён особенностями занимаемой им экологической ниши, накладывающей меньшие ограничения на размеры генома. Существует мнение, что эти особенности позволили мимивирусу накопить большое количество копий гомологичных генов, которые возникали в результате дупликации генов и их дальнейшей независимой эволюции[15].
Вопрос о происхождении гигантских вирусов остаётся ещё более загадочным, чем вопрос их эволюции. Было высказано предположение, что представители группы крупных ядерно-цитоплазматических ДНК-содержащих вирусов (поксвирусы, иридовирусы, фикоднавирусы, мимивирусы и др.) ведут своё начало от более сложных (возможно клеточных) форм, таких как современные микоплазмы и риккетсии[48]. В пользу этой гипотезы говорит наличие в геноме крупных ДНК-содержащих вирусов большого количества «избыточных» генов, которые не являются строго необходимыми для размножения и функционально дублируют хозяйские. Сторонники этой точки зрения рассматривают, например, наличие у мимивируса и Megavirus chilensis неполного аппарата трансляции как признак того, что они произошли от свободноживущего или паразитического предка, у которого этот аппарат был полностью функционален[4][24]. По мере открытия новых гигантских вирусов учёные надеются получить больше информации о возможном общем предке крупных ядерно-цитоплазматических ДНК-содержащих вирусов.
Альтернативная гипотеза вирусного эукариогенеза, напротив, предполагает возникновение ядра эукариотических клеток из крупных ДНК-содержащих вирусов, подобных мимивирусам[48]. На разных этапах этого процесса новое примитивное ядро предположительно могло несколько раз возвращаться к существованию в форме гигантского вируса, что привело бы к формированию нескольких независимых групп вирусов.
В 2000 году Международный комитет по таксономии вирусов официально объявил, что вирусы не относятся к живым организмам. Однако с открытием мимивируса эта идея была снова поставлена под сомнение[49][50]. Хотя мимивирус, как и любой другой вирус, не имеет генов рибосомальных белков и использует рибосомы хозяина, он формирует вирусные фабрики, которые функционируют относительно независимо от клетки. Было высказано мнение, что именно вирусные фабрики, а не метаболически неактивные вирусные частицы, и следует рассматривать как вирус[48]. Автор этой идеи полагает, что вирусная фабрика, осуществляющая репликацию ДНК, транскрипцию генов и синтез соответствующих белков с привлечением компонентов цитозоля, очень сильно напоминает клеточное ядро. С этой точки зрения мимивирус кажется гораздо ближе к живому, чем его вирион.
Тем не менее, многие учёные продолжают придерживаться более традиционного в последнее десятилетие мнения о том, что вирусы представляют собой неживые частицы[51]. Возможно, более точно на этот вопрос можно будет ответить с открытием новых гигантских вирусов.
Мимивирус имеет примерно икосаэдрический капсид диаметром 400—500 нм. Капсид покрыт многочисленными белковыми нитями длиной 80—120 нм. В научной литературе приводятся размеры вириона от 400 до 800 нм, в зависимости от того, замеряется ли диаметр капсида или общая длина вируса вдоль продольной оси, включая белковые нити. Основной белок капсида мимивируса является продуктом гена L425 и состоит из двух доменов с укладкой типа jelly-roll («рулет»). Этот белок формирует гомотримерные капсомеры — единицы организации капсида. Капсомеры упакованы гексагонально в виде «ромашек»: шесть капсомеров окружают одно углубление между ними. Также в составе капсида обнаруживается структурный белок сердцевины L410.
На одной из вершин капсида обнаружена звёздчатая структура, лучи которой разделяют пять треугольных граней, сходящихся в этой вершине. Лучи имеют ширину около 50 нм, толщину — 40 нм и длину — 200 нм, почти доходя до соседних вершин. Присутствие этой структуры изменяет взаимное расположение граней вириона, в результате чего его форма отклоняется от идеально икосаэдрической: через вирион можно провести только одну ось пятилучевой симметрии, которая проходит через вершину, отмеченную звёздчатой структурой. Так как на поверхности звёздчатой структуры не наблюдается гексагонально упорядоченных углублений, предполагают, что она состоит из белка, который отличается от основного белка капсида. Данная структура играет особую роль при заражении клетки-хозяина: при инфицировании звёздчатая «застёжка» открывается, и через это место ДНК вируса выходит из капсида. По этой причине звёздчатую структуру ещё называют «звёздными вратами».
У мимивируса не наблюдается внешней оболочки, что указывает на то, что он покидает заражённую клетку не путём экзоцитоза.
Капсид мимивируса покрыт снаружи густым слоем длинных белковых нитей. Исследование этих нитей с помощью атомно-силового микроскопа показало, что они часто бывают присоединены к общей несущей структуре и заканчиваются небольшой глобулой. Однако до сих пор не известно, к каким участкам поверхности капсида они прикрепляются. Белковые нити устойчивы к воздействию протеаз до тех пор, пока их не обработать лизоцимом, это указывает на то, что эти нити покрыты пептидогликаном. Это хорошо согласуется с тем, что мимивирус окрашивается по методу Грама. Существует мнение, что сильно гликозилированная поверхность нитей может играть роль в привлечении амёб-хозяев.
НуклеокапсидМимивирус обладает многими особенностями строения, характерными для других крупных ядерно-цитоплазматических ДНК-содержащих вирусов. Например, сразу под капсидом у мимивируса находятся два электронноплотных слоя, которые предположительно являются мембранами. Под этими мембранами находится белковая оболочка толщиной около 7 нм, внутри которой заключена линейная двухцепочечная ДНК вируса. Все описанные компоненты образуют так называемый нуклеокапсид. Стенки нуклеокапсида отстают от стенок капсида примерно на 30 нм, в районе звёздчатой структуры поверхность нуклеокапсида вдавлена. Предполагают, что пространство между вершиной звёздчатой структуры и нуклеокапсидом может быть заполнено гидролитическими ферментами, которые нужны для проникновения вируса в клетку. Между капсидом и нуклеокапсидом были обнаружены внутренние белковые нити, которые предположительно обеспечивают стабильное положение второго внутри первого.
Неструктурные белки и РНКПомимо структурных белков капсида, в составе вириона обнаруживают и другие белки, которые классифицируют на несколько функциональных групп:
белки, участвующие в транскрипции, 5 субъединиц ДНК-зависимой РНК-полимеразы, 2 хеликазы (R350, L540), кэпирующий фермент, 4 фактора транскрипции (L377, L538, L544, R563), белки окислительных путей (предположительно помогают вирусу преодолеть окислительный стресс, связанный с активацией защитных систем клетки-хозяина) (9), белки, модифицирующие липиды и белки, 2 протеинкиназы, протеинфосфатаза, фосфоэстераза, липаза, белки, участвующие в метаболизме ДНК, топоизомеразы IA и IB, эндонуклеаза, корректирующая повреждения ДНК ультрафиолетовым светом.Кроме белков и ДНК, из очищенных вирионов были выделены несколько различных мРНК, кодирующих ДНК-полимеразу (R322), главный белок капсида L425, TFII-подобный фактор транскрипции (R339), 3 аминоацил-тРНК-синтетазы (L124, L1164 и R663) и 4 неидентифицированных белка, специфичных для мимивируса. По-видимому, трансляция этих мРНК клеточным аппаратом биосинтеза белка необходима для начала репликации вируса. Другие ДНК-содержащие вирусы, такие как цитомегаловирус (Cytomegalovirus) и вирус простого герпеса (Herpes simplex virus type-1), также содержат мРНК.
{{Delete|G13}}提交刪除。 种 Acanthamoeba polyphaga mimivirus
拟菌病毒是一个包括Acanthamoeba polyphaga mimivirus(APMV)的一个属,或许是与演化史相关的巨型病毒。[1]通常所说的“拟菌病毒”就指APMV。在口語中,APMV更普遍簡稱為“米米病毒”(mimivirus)。2011年10月中旬,科學家發現了Megavirus chilensis,這是已知衣壳直径最大的病毒。[2]拟菌病毒比起其他病毒有着庞大的基因组。拟菌病毒(mimicking microbe)的命名反映了它的体积及革兰氏染色的特性。[3]
拟菌病毒最初是在1992年的一次对军团病研究中,偶然在变形虫多食棘阿米巴(Acanthamoeba polyphaga)中发现的。然而在之后的革兰氏染色实验中被错误地认为是一种革兰氏阳性菌,并被命名为“布拉德福德球菌”(Bradfordcoccus)。到2003年,法国马赛地中海大学在《科学》期刊上发布了文章,认定这个有机体为一种病毒。[4]
国际病毒分类委员会将拟菌病毒归为拟菌病毒科。[5]并在生物病毒分类中归为I类病毒(双链DNA病毒)。[6]在不严格的分类下,拟菌病毒加入核质巨DNA病毒(NCLDV)一类。这类病毒都有着类似的分子特性和庞大的基因组。拟菌病毒同样有21个同源于NCLDV的高度保守基因,并且进一步的研究也显示拟菌病毒可能是NCLDV的一个分支。[4]
拟菌病毒的衣壳直径达400nm, 是仅次于潘多拉病毒和近期发现的Megavirus chilensis的第三大病毒。加上衣壳表面的蛋白丝,总长度可达600nm. 各种文献对此的描述高度相似,病毒大小都在400nm到800nm之间。在电子显微镜下,病毒的衣壳呈六边形,因此衣壳的立体结构为正二十面體。[7]拟菌病毒并没有包膜,这意味着这类病毒并非通过胞吐被放出细胞的。[8]
发现拟菌病毒的小组后来发现了一种略微更大的病毒,命名为妈妈病毒(mamavirus) ,以及能感染它的亚病毒Sputnik virophage.[9]
拟菌病毒有着所有NCLDV上共同的形态学特征。M. Suzan-Monti等人提出,如同其他NCLDV病毒,拟菌病毒可能也有这内含的包裹DNA核的脂质层。在电子显微镜下,病毒体浓缩的核呈现出黑色。病毒庞大的基因组就在这个区域之中。
一些mRNA的转录子可以被拟菌病毒独立完成。如同其他NCLDV的DNA聚合酶的转录,一种衣壳蛋白质和一个类似于TFII的转录因子也被发现。然而,三种明显不同的氨酰-tRNA合成酶的转录子以及4种未知的拟菌病毒独有的mRNA分子也被发现。这些预先打包的转录子能够在病毒基因没有表达的情况下被翻译,并且可能对拟菌病毒的复制是必要的。其他的DNA病毒,如人类巨细胞病毒和单纯疱疹病毒I型也有mRNA预先打包转录的特点。[8]
拟菌病毒的基因组是一个线性的,有着1,181,404对碱基的DNA双链。[10]是目前已知病毒中基因组最庞大的一种,超出了仅次于它的伦比约餐厅虫病毒(Cafeteria roenbergensis virus)450,000对碱基。此外,它甚至比至少30种细胞体还大。[11]
除了庞大的基因组,拟菌病毒还有着大约979个蛋白质编码基因,远远超出了病毒最低所需的4个基因(参见 MS2噬菌体和Qβ噬菌体)。[12]分析显示,拟菌病毒特有的基因包括氨酰tRNA合成酶以及其他一些早前被认为只能由细胞微生物编码的酶。正如其他的巨型DNA病毒,拟菌病毒也被发现有着其他病毒不具有的与糖类、脂类和氨基酸代谢相关的基因。[8]大约90%的基因组是编码基因,而另外10%为非编码DNA。
拟菌病毒的复制过程尚不清楚。但是我们至少知道此病毒能与阿米巴细胞膜上的一种受体结合。一旦进入阿米巴,便进入一段隐蚀期,这时病毒会消失,细胞会正常工作。在感染约4h后,细胞表面会显现出一些聚集物。感染8h后,在细胞中的拟菌病毒便清晰可见。细胞质会持续被新组装的病毒体填充。感染约24h后,细胞便极为可能破裂,并释放出新的拟菌病毒病毒体。[8]
目前我们对拟菌病毒的复制周期几乎一无所知,显然是附着于细胞表面,进入,病毒核物质释放,DNA复制,转录,翻译,组装,最后释放子病毒体。然而,研究人员是使用对被感染的细胞的显微照相来得出上述综述的。这些显微照片显示了拟菌病毒的衣壳在细胞核中组装,并在出核时获得一个脂质膜,以及一些与其他包括NCLDV的不少病毒相似的颗粒。这些颗粒在其他病毒中被称为病毒工厂(viral factories ),它能使宿主细胞被更多地改造,并病毒的组装更快。
拟菌病毒或许是某些肺炎的病原体。这种推测是基于在某些肺炎患者体内发现此病毒抗体的间接证据。[13]然而,将拟菌病毒归为病原体的理论是极为脆弱的,因为仅有极少的几篇文章报告了拟菌病毒与肺炎的潜在关系。造成肺炎的最大因素目前还尚未知晓。[14]即便如此,已经有一种拟菌病毒从一名突尼斯的女性肺炎患者中分离出来。[15]
拟菌病毒的许多特性将之放在了生命与非生命之间。它并不比某些细菌小,如康氏立克次氏体(Rickettsia conorii)和Tropheryma whipplei,并且有着与不少细菌相当甚至更多的基因。在之前病毒还没有被认为能够进行编码。此外,拟菌病毒还有着核苷酸和氨基酸合成相关的基因,这甚至是一些小型的专性寄生细菌所不具有的。然而,拟菌病毒确实缺少了核糖體蛋白質,这使得他们必须依靠宿主细胞的蛋白转运和能量代谢。[來源請求]
因为拟菌病毒的起源非常古老,甚至有可能先于细胞[16][17],拟菌病毒已经在生命起源的争论中成为一种论据。一些拟菌病毒独有的基因,包括那些衣壳相关的基因,在多种能感染各域生物的病毒中保留。这暗示了拟菌病毒与一种先于细胞生物并且在地球生物的起源中起重要作用的DNA病毒有联系。[16]另一种假设是,有明显的三种DNA病毒与目前的三个域(真核生物、古生菌、细菌)的产生有关。[17]也有猜想认为这些病毒可能一开始是细菌,后来退化成了病毒。[來源請求]
然而拟菌病毒并未表现出以下特征:具有稳态,能量代谢,能对刺激作出反应,自我繁殖,具有细胞结构等通常用以定义生命的特征。
|date=中的日期值 (帮助) 拟菌病毒是一个包括Acanthamoeba polyphaga mimivirus(APMV)的一个属,或许是与演化史相关的巨型病毒。通常所说的“拟菌病毒”就指APMV。在口語中,APMV更普遍簡稱為“米米病毒”(mimivirus)。2011年10月中旬,科學家發現了Megavirus chilensis,這是已知衣壳直径最大的病毒。拟菌病毒比起其他病毒有着庞大的基因组。拟菌病毒(mimicking microbe)的命名反映了它的体积及革兰氏染色的特性。

ミミウイルス (Acanthamoeba polyphaga mimivirus) は、1992年にアカントアメーバの中で発見されたウイルスである。2011年10月中旬にチリでメガウイルス・キレンシス (Megavirus chilensis) を発見するまでは知られるウイルスの中で最も巨大であった[1]。
本種のみでミミウイルス属を構成する。上位のミミウイルス科には複数の種が発見されており、近縁の種にママウイルス (Mamavirus) が存在する。グラム陽性菌に「化けて (mimick)」いたという意味で命名された[2]。
ミミウイルスは、直径がおよそ750nmあり、20面体のカプシド内に脂質二重膜で覆われたDNAがあり、このカプシドを毛状構造が覆う形状を持つ [3]。
また、ゲノムサイズはおよそ120万塩基対、遺伝子数は980あり[4]、巨大核質DNAウイルスの一つに数えられる。
1992年、イングランドのウェスト・ヨークシャー州ブラッドフォードにある病院の冷却水から、肺炎の原因菌を調査中に発見された[5][6]。この時は、ブラッドフォード球菌と名付けられたが、その後の調査によりリボソームRNAを持たないことが判明、2003年にサイエンス誌に発表された論文において、ミミウイルス(Mimivirus)と名付けられた [7]。
ミミウイルス (Acanthamoeba polyphaga mimivirus) は、1992年にアカントアメーバの中で発見されたウイルスである。2011年10月中旬にチリでメガウイルス・キレンシス (Megavirus chilensis) を発見するまでは知られるウイルスの中で最も巨大であった。
本種のみでミミウイルス属を構成する。上位のミミウイルス科には複数の種が発見されており、近縁の種にママウイルス (Mamavirus) が存在する。グラム陽性菌に「化けて (mimick)」いたという意味で命名された。
미미바이러스(Mimivirus)는 바이러스의 일종으로, 미미바이러스과에 속한다. 또한, 이것은 세포생리학적으로 관련된 큰 바이러스의 집단을 가리킨다. 구어체로, APMV는 흔히 미미바이러스라고 불리기도 한다. 모방 미생물의 줄임말인 미미바이러스는 바이러스의 큰 크기와 그람 자국의 성질을 반영하여 그렇게 불린다. 또한, 미미바이러스는 다른 바이러스들보다 더 복잡한 유전자를 가지고 있다. 더 큰 바이러스인 판도라바이러스가 설명된 2013년까지 알려진 바이러스 들 중 캡시드 지름이 가장 컸다.
APMV는 1992년 마르세유와 리즈의 연구자들이 레지오넬라증을 연구하던 도중에 아메바 아칸타아메바 폴리파가에서 우연히 발견되었다. 이 바이러스는 그람 염색을 통해 관찰되었고, 그람 양성 박테리아로 잘못 생각되었다. 결과적으로 이것은 아메바가 발견된 영국의 브래드퍼드 대학교의 이름을 따 브래드퍼드코커스라고 명명되었다. 2003년에 프랑스 마르세유에 있는 유니버스테 데 라 메디테라네(Université de la Méditerranée)의 연구원들이 미생물을 바이러스로 인식하는 논문을 사이언스지에 발포하였다. 미미바이러스는 그람 염색의 박테리아의 모습을 닮아서 "미생물을 닮았다"는 뜻의 미미바이러스라고 명명되었다. 미미바이러스를 발견한 연구 팀은 나중에 마마바이러스로 불리는 조금 더 큰 바이러스와 그것을 감염시키는 스푸트니크 바이러스도 발견하였다.
미미바이러스 국제 바이러스 분류 위원회에 의해 미미바이러스과의 일원으로 바이러스계에 편입되어 볼티모어 분류 체계 I조에 편입되었다. 엄격하게 분류하는 방법은 아니지만 미미바이러스는 핵세포질 대 바이러스(NCLDV)로 알려진 거대 바이러스 그룹에 속하며 모두 분자의 특성과 큰 유전자들을 공유한다. 또한, 미미바이러스 유전자는 대부분의 NCLDV에서 보존도가 높은 그룹의 초기 분립자임이 밝혀졌다.
이 미미바이러스는 메가바이러스 칠렌시스, 판도라바이러스, 피토바이러스에 이어서 4번째로 큰 바이러스이다. 미미바이러스의 캡시드 직경은 400 nm 정도이다. 캡시드 표면에서 100 nm 크기의 단백질 필라멘트는 바이러스의 총길이를 600nm까지 키운다. 과학 문헌의 발달은 이러한 수치를 매우 가깝게 제시하는데 전체 길이 또는 캡시드 직경이 실제로 인정되는지 여부에 따라서 바이러스의 총 길이가 400 nm에서 800 nm 사이로 에상된다. 그것의 캡시드는 전자 현미경아래에 육각형으로 나타나기 때문에 캡시드 대칭은 20면체이다. 또한, 외부 바이러스 막을 가지고 있는 것처럼 보이지 않으며, 바이러스가 난세포에 의해 숙주 세포에서 빠져나가지 않는다는 것을 알 수 있다.
미미바이러스는 NCLDV 바이러스 그룹의 구성원과 몇 가지의 형태학적 특성들을 공유한다. 비리온의 응축된 중심핵은 전자 현미경에서 어두운 영역으로 나타나며 바이러스의 큰 유전자들은 대부분 이 지역에 서식한다. 중심핵을 둘러싸고 있는 내부의 지질층이 다른 모든 NCLDV 바이러스에도 존재하기 때문에 이 특징 또한 미미바이러스에도 존재할 수 있다. 또한, 정제된 바이러스로부터 여러 mRNA 정보를 복구할 수 있다. 다른 NCLDV와 마찬가지로 DNA 중합효소, 캡시드 단백질, TFII 유사 전사계수의 정보가 발견되었다. 그러나 3개의 구별되는 아미노아실 tRNA 합성 효소 정보와 미미바이러스에 특유한 4개의 알려지지 않은 mRNA 분자도 발견되었다. 미리 포장된 이 정보들은 바이러스의 유전자 발현 없이도 번식이 가능하며 복제에 미미바이러스가 필요할 가능성이 높다. 인간 사이토메갈로 바이러스, 헤르페스 심플렉스 바이러스-1도 미리 포장된 mRNA 기록이 특징이다.









_Niemeyer_virus.jpg)
.png)
.png)


















{kind=link}