
These small aquatic crustaceans are very diverse and are the most numerous metazoans in the water community...
The World Association of Copepodologists (WAC) is a nonprofit international organization whose purpose is to promote research on Copepoda by facilitating communication among interested specialists...
The copepods include around 12,000 described species of mostly small aquatic crustaceans. Most are between 0.5 mm and 10 mm in length, but some free-living forms exceeed 1.5 cm and some parasitic forms may reach 25 cm. Copepods are extremely abundant in the ocean (from the surface to 5000 meters deep, including at least one species described from a hydrothermal vent), in freshwater, in estuarine habitats, and in interstitial habitats (i.e, between sand grains). The majority of known free-living copepods belong to the Calanoida, Harpacticoida, or Cyclopoida, although these groups also include some parasitic species. Calanoids have greatly elongated antennules. Most calanoids are planktonic and as a group they are very important primary consumers in both freshwater and marine food webs. Harpacticoids have a more worm-like shape, with (in contrast to both calanoids and cyclopoids) the posterior segments not much narrower than the anterior ones. Both the antennules and antennae are quite short in harparcticoids; cyclopoids have moderately long antennae, although never as long as the antennules of calanoids. The antennae are uniramous (i.e., unbranched) in cyclopoids, but biramous in calanoids and harpacticoids. Most harpacticoids are benthic (bottom-dwelling) and occur in a wide range of aquatic environments; at least a few freshwater and marine species are known to form cysts. Cyclopoids are found in both marine and freshwater habitats and most are planktonic. (Brusca and Brusca 2003)
The non-parasitic copepods move by crawling or swimming, using some or all of the thoracic limbs. Many of the planktonic forms have dense setae on their appendages, making them resistant to sinking. Calanoids are mainly planktonic feeders. Although benthic harpacticoids are often reported to be detritus feeders, many feed mainly on microorganisms living on the surface of detritus or sediment particles (e.g., diatoms, bacteria, and protists). (Brusca and Briusca 2003; Margulis and Chapman 2010)
Of the seven remaining orders, the Mormonilloida are planktonic; the Misophrioida are known from deep sea epibenthic habitats as well as anchialine caves in both the Atlantic and Pacific; and the Monstrilloida are planktonic as adults, but as larvae are endoparasites of gastropod mollusks, polychaete annelid worms, and occasionally echinoderms. Members of the orders Poecilostomatoida and Siphonostomatoida are exclusively parasitic. Siphonostomatoids are ectoparasites or endoparasites of various invertebrates as well as marine and freshwater fishes. They are often very tiny and may exhibit a reduction or loss of body segmentation. Poecilostomatoids parasitize invertebrates and marine fishes and may also show a reduced number of body segments. The Platycopioida are benthic copepods known mainly from marine caves. The Gelyelloida are known only from European groundwaters. (Brusca and Brusca 2003; Margulis and Chapman 2010)
Els copèpodes, del grec 'potes de rem', (Copepoda), són crustacis de mida molt petita, generalment microscòpics, que es troben abundantment, tant en aigua dolça com salada.
Aquest grup de crustacis no tenen closca, i el seu desenvolupament s'inicia a partir d'una larva naupli. El copèpode d'aigua dolça més estudiat és Cyclops que ofereix un quadre general del grup bastant representatiu. El seu aspecte és el d'un crustaci decàpode, amb una porció anterior amb forma de pera que inclou la regió cefàlica i els dos primers segments del tòrax. A continuació es troben tres segments del tòrax mòbils, i finalment l'abdomen, sense apèndixs i que acaba en una cua en forma de forca, amb filaments plomosos. Tenen un sol ull, que és una evolució dels dos ulls fusionats, i les antenes principals, molt articulades són molt més grans que les secundàries. Posseeixen 4 parells d'apèndixs toràcics que els serveixen per nedar i vestigis d'un cinquè parell que en altres espècies de copèpodes són més evidents. A més de les mandíbules i maxil·les, el primer parell d'apèndixs toràcics ha evolucionat fins a convertir-se en maxil·les secundàries.
Dins dels Copèpodes trobem dos subgrups principals:
Molts eucopèpodes són de cos transparent; alguns tenen una certa coloració, com és el cas dels Anomalocera. Com fet curiós, es pot notar que algunes espècies que són translúcides arriben a acolorir abundantment l'aigua a causa de la difracció de la llum en passar pels seus cossos. Algunes espècies marines són lluminoses per si mateixes, ja que tenen en el tegument una substància fosforescent.
Els copèpodes són tan abundants en l'aigua de mar, que en qualsevol mostra de plàncton n'apareixen exemplars. Com la gran majoria dels copèpodes pertanyen al subgrup dels eucopèpodes, trobem exemplars distribuïts al llarg de totes les latituds, fins i tot a l'Antàrtida.
Moltes de les espècies compten amb apèndixs plomosos que en molts casos poden arribar a superar la mida de l'animal, fins i tot en 4 o 6 vegades la seva llargària. En aigua dolça també els podem trobar per molts llocs, en una distribució tan ampla com la dels Cladòcers.
Els animals pertanyents a aquest subgrup de copèpodes, viuen totes en aigua dolça i pertanyen tots a la família dels Argúlids, composta dels gèneres Argulos, Dolops i Chonopeltis. La gran majoria són paràsits, encara que es poden trobar algunes espècies nedant en llibertat. A diferència dels eucopèpodes, a més de l'únic ull fusionat, posseeixen dos ulls simples als costats del cos. El cos dels branquiurs és translúcid, fins al punt que únicament s'aprecia la zona abdominal on es troba l'aparell digestiu.
La gran majoria dels copèpodes neden lliurement però existeixen algunes espècies que s'han convertit en paràsites. Moltes d'aquestes últimes conserven un aspecte similar al de les formes lliures mentre d'altres han arribat a l'extrem de perdre les seves característiques de crustaci i fins i tot d'artròpode, observant-se molt més en el cas de les femelles. Per poder classificar aquestes últimes espècies dins dels copèpodes, es parteix de l'estudi de mascles joves, els quals no estan gaire modificats.
Dins dels eucopèpodes hi ha una gran quantitat d'espècies paràsites, més que en cap altre grup de crustacis. Són molt coneguts els anomenats "polls dels peixos" de la família dels calígids, que es caracteritzen pel cos ample i deprimit que els facilita adherir-se al seu hoste fàcilment. Una particularitat dels calígids, és que en les seves primeres etapes de desenvolupament neden lliurement, i només inicien la seva etapa parasitària quan són adults joves. Romanen paràsits sense facultats natatòries durant un període determinat per etapes de creixement i aparellament i finalment retornen a espècimens lliures, recuperant les seves capacitats natatòries.
Un altre paràsit de costums curiosos és el Lernacea, les larves del qual que neixen en forma de naupli, inicien la seva vida paràsita sobre un peix de la família dels pleuronèctids, com un llenguado; els seus apèndixs es redueixen fins a quedar convertits en mers monyons. Quan arriba l'època d'aparellament, l'animal es desprèn de l'hoste. Després d'acoblar-se els mascles moren, i les femelles busquen un nou hoste, generalment un bacallà o un lluç, es fixen a les brànquies i inicien un procés degeneratiu i arriben a perdre totalment els apèndixs fins a quedar convertides en sacs, sense cap aparença de crustaci.
Com ja s'ha comentat, la major part dels branquiurs són paràsits. Els paràsits solen fixar-se sobre les ganyes del peix, de vegades en quantitats tan grans que poden produir la mort de l'animal. La boca s'atura fins a convertir-se en un simple conducte xuclador, en tant que els apèndixs es redueixen, encara que no arriben a desaparèixer del tot. A diferència d'altres copèpodes paràsits, els branquiurs no recuperen la seva forma lliure una vegada han iniciat la seva etapa parasitària, sinó fins i tot el final de la seva vida en l'època d'aparellament.
Els copèpodes, del grec 'potes de rem', (Copepoda), són crustacis de mida molt petita, generalment microscòpics, que es troben abundantment, tant en aigua dolça com salada.
Klanonožci (Copepoda) je skupina korýšů řazená obvykle jako podtřída do třídy Maxillopoda. Jsou to obvykle rozměrově menší (1–2 mm) vodní organizmy, tvořící často významnou součást planktonu ve slaných i sladkých vodách.[1] Mají dlouhé antenuly a anteny, hlavu často srůstající s přilehlými hrudními články, naupliové očko, po stranách těla samiček se nalézají vaječné váčky.[2]
Jsou to filtrátoři, dravci nebo i parazité. Ke klanonožcům patří vznášivky (Gymnoplea), buchanky (Cyclopoida), plazivky (Harpacticoida) a další drobní korýši.[2] V ČR je známo asi 220 druhů.[1]
Klanonožci (Copepoda) je skupina korýšů řazená obvykle jako podtřída do třídy Maxillopoda. Jsou to obvykle rozměrově menší (1–2 mm) vodní organizmy, tvořící často významnou součást planktonu ve slaných i sladkých vodách. Mají dlouhé antenuly a anteny, hlavu často srůstající s přilehlými hrudními články, naupliové očko, po stranách těla samiček se nalézají vaječné váčky.
Jsou to filtrátoři, dravci nebo i parazité. Ke klanonožcům patří vznášivky (Gymnoplea), buchanky (Cyclopoida), plazivky (Harpacticoida) a další drobní korýši. V ČR je známo asi 220 druhů.
Vandlopper (Copepoda) er en gruppe af krebsdyr, der forekommer i både ferskvand og saltvand. Nogle arter lever frit i vandet som plankton, andre lever på havbunden og nogle lever fugtige steder på landjorden, f.eks. under visne blade i skovbunden eller mellem fugtigt mos. Nogle arter lever som snyltere, fortrinsvis på fisk, men er fundet på næsten alle slags dyr i havet og i ferskvand.[1] Der findes omkring 13.000 arter af vandlopper, hvoraf 2.800 lever i ferskvand.[2] De deles i 10 ordener.
Vandlopper er et meget vigtigt led i fødekæden i havet, hvor de optræder i meget store mængder på grund af deres hurtige vækst og mange årlige generationer. De er meget effektive til at filtrere mikroskopiske planktonalger og er det vigtigste fødeemne for fiskeynglen. Også visse arter af voksne fisk og hvaler, f.eks. sild, makrel og sejhvalen, lever især af vandlopper.[1]
Kropslængden er normalt mellem 0,2 og 2 millimeter. Den mindste art, Sphaeronellopsis monothrix, er 0,11 mm lang og lever i muslingekrebs (Ostracoda). Den største art, Pennella balaenoptera kan blive 32 centimeter lang og lever som snylter i bardehvaler, hvor den sætter sig fast i muskulaturen under spæklaget.
Vandlopper kan muligvis i fremtiden indirekte blive truet af store mængder CO2, som opsuges i verdenshavene og dermed forsurer vandet og gør det mindre basisk, viser forskning fra Utah State University i 2017. En forsker udtaler, at "hvis der sker noget med vandlopper, påvirkes alt det, der er afhængig af dem, og det er stort set alt". Vandlopper er bl.a. byttedyr for de større fiskearter. Når havene bliver mere sure, får nogle af vandloppernes største fjender bedre levevilkår. Dette gælder ifølge forskerne for nogle gopler kaldet havhvepse, der i et eksperiment fik bestanden af vandlopper fra Australien til at mindskes med 83% i en vandtank, hvor vandet var blevet kunstigt forsuret til det niveau der forventes for år 2100 samtidig med at man havde tilsat havhvepse. Hvis dette også er tilfældet for vandlopper i arktiske områder, hvor de er særligt udbredt, vil det blive "virkeligt alvorligt".[3]
|chapter= ignoreret (hjælp) Vandlopper (Copepoda) er en gruppe af krebsdyr, der forekommer i både ferskvand og saltvand. Nogle arter lever frit i vandet som plankton, andre lever på havbunden og nogle lever fugtige steder på landjorden, f.eks. under visne blade i skovbunden eller mellem fugtigt mos. Nogle arter lever som snyltere, fortrinsvis på fisk, men er fundet på næsten alle slags dyr i havet og i ferskvand. Der findes omkring 13.000 arter af vandlopper, hvoraf 2.800 lever i ferskvand. De deles i 10 ordener.
Vandlopper er et meget vigtigt led i fødekæden i havet, hvor de optræder i meget store mængder på grund af deres hurtige vækst og mange årlige generationer. De er meget effektive til at filtrere mikroskopiske planktonalger og er det vigtigste fødeemne for fiskeynglen. Også visse arter af voksne fisk og hvaler, f.eks. sild, makrel og sejhvalen, lever især af vandlopper.
Ruderfußkrebse (Copepoda) sind kleine Krebse (Crustacea), die marine und limnische Gewässer besiedeln. Sie bilden mit rund 13.000 Morphospezies die artenreichste Gruppe der Crustaceen und stellen den größten Anteil des marinen Zooplanktons dar. Sie sind daher ein sehr wichtiges Element der marinen Nahrungskette. Sie leben als Filtrierer, Räuber oder Parasiten. Bei Letzteren kann aufgrund der Lebensweise eine stark modifizierte Morphologie auftreten.
Die Körperlänge beträgt überwiegend zwischen 0,2 und 2 mm. Die kleinste Art, Sphaeronellopsis monothrix (Männchen 0,11 mm), lebt im Brutraum von Muschelkrebsen (Ostracoda). Die größte Art, Pennella balaenoptera, ein Parasit an Bartenwalen, erreicht eine Körperlänge von 320 mm. Die ursprüngliche Körpergestalt ist langgestreckt bis tropfenförmig, ist jedoch innerhalb der Gruppe je nach Lebensweise stark abgewandelt. Pelagische Arten entsprechen noch dem ursprünglichen Bauplan. Bodenbewohner sind mehr abgeflacht und im Substrat wühlende Arten eher zylindrisch gebaut. Am meisten modifiziert sind parasitische Arten, bei denen die adulten Weibchen kaum mehr als Copepoda zu erkennen sind und deren Zugehörigkeit zu den Ruderfußkrebsen nur aufgrund der Entwicklungsstadien ermittelt werden kann.
Der Körper setzt sich aus dem Kopf, zehn Rumpfsegmenten und dem Telson zusammen. Kopf und 1. Thorakalsegment sind stets verschmolzen. Ein Carapax fehlt. Der Rumpf gliedert sich in sechs Thoraxsegmente mit Gliedmaßen und vier Abdominalsegmente, die über das Hauptkörpergelenk verbunden sind. Die 1. Thorakopoden sind zu einästigen Maxillipeden umgewandelt. Die folgenden 4 Paare sind zweiästige Schwimmbeine. Das sechste und letzte Paar ist oft reduziert oder einästig. Die Geschlechtsöffnung befindet sich am 1. Abdominalsegment, wo beim Weibchen ein oder zwei Eisäcke befestigt sind. Entsprechend der Lebensweise ist auch die Antennula gestaltet. Bei den Calanoida ist sie bedeutend länger als, bei Cyclopoida hingegen kürzer als halbe Körperlänge. Die am Telson stehenden Furcaläste dienen als Schwebefortsätze bzw. Steuerorgane. Die Gestalt der Mandibeln und des Mundvorraums spiegeln die Lebensweise wider und sind wichtige taxonomische Merkmale. So ist bei den Siphonostomatoida Labrum und Labium zu einem Kegel verschmolzen und manchmal in ein langes Saugrohr ausgezogen. Die Färbung der Copepoda beruht auf eingelagerten Öltröpfchen, die als Reservefett dienen. Nur selten spielen eigene Hautpigmente eine Rolle.
Die Copepoda sind kosmopolitisch verbreitet und besiedeln sämtliche Gewässertypen der Erde, von den Ozeanen bis zum Kleinstgewässer. Sie fehlen nur in den von Dauereis bedeckten und in den wasserlosen Gebieten. Ihr Verbreitungsschwerpunkt liegt mit rund 80 % der rezenten Spezies im Meer, wo sie mit der größten Individuen- und Artenzahl in den oberen Schichten und im Litoral vorkommen. Copepoda wurden bis in eine Tiefe von 5000 m nachgewiesen. Die besiedelten Lebensräume sind äußerst vielfältig. Prinzipiell kann zwischen frei schwimmenden, am Boden lebenden, im Substrat wühlenden und parasitären Arten unterschieden werden. An Land werden auch feuchte Moose und Blattachseln besiedelt. Einige Arten sind auch ins Grundwasser eingedrungen.
Insbesondere sehr alte Seen sind Hotspots der Biodiversität. Von den 120 im Baikalsee vorkommenden Spezies sind die meisten endemisch.[1] Ähnliches gilt für den Tanganjikasee.
Die Furchung der Eier ist in der Regel holoblastisch zu gleichen Teilen (total-adäqual), manchmal auch zu ungleichen Teilen (inadäqual). Einige Parasiten scheinen rein superfiziell zu furchen. Aus dem Ei schlüpft fast stets eine Naupliuslarve. Verschiedene Parasiten bilden eine Ausnahme, da sie einen verkürzten Entwicklungszyklus aufweisen und bereits eine Larve im Copepoditstadium schlüpft. Die Jungtiere durchleben in der Regel sechs Stadien als Naupliuslarven, wobei es in jedem Stadium zur Häutung kommt. Auch in den sechs Copepoditstadien häuten sich die Tiere weiter, wobei sich die Zahl der Segmente und Beinpaare vermehrt. Das 12. Stadium, also das sechste Copepoditstadium, ist der Adultus, der sich nicht mehr häutet. Die Lebensdauer ist stark von der Temperatur abhängig. Die Lebensdauer des Muschelparasiten Mytilicola intestinalis beträgt im Mittelmeer 3–6 Monate, wobei sich innerhalb eines Jahres mehrere überlappende Generationen bilden. In den kühleren Gewässern der englischen Südwestküste treten nur zwei Generationen während des Sommers auf. Die Individuen hier erreichen ein Lebensalter von 9–12 Monaten. Für größere Arten wie Eudiaptomus vulgaris werden 10–13 Monate, für Megacyclops viridis 8–14 Monate angegeben. Als Überlebensstrategie während ungünstiger Jahreszeiten, vor allem im Winter, bilden viele Arten Dauereier. Wenige Arten bilden als Adulte jedoch Zysten und sind vor allem im Winter aktiv. Sie überdauern den Sommer in den Zysten.
Alle Copepoda sind getrennt geschlechtig und weisen einen ausgeprägten Sexualdimorphismus auf. Die Männchen sind stets kleiner als die Weibchen und die Antennula ist als Greiforgan ausgebildet. Auch die übrigen Kopfgliedmaßen und Thorakopoden können unterschiedlich ausgeprägt sein. Das Geschlechterverhältnis ist bei vielen Arten deutlich zu Gunsten der Weibchen verschoben. Bei manchen Arten konnten bis heute überhaupt keine Männchen gefunden werden, was vermutlich an dem raschen Absterben nach der Befruchtung liegt. Parthenogenese tritt nur bei wenigen Arten auf. Die Begattung ist bei den Calanoida gut untersucht. Das Männchen greift mit seiner geknieten Antenne (meist die rechte) das Weibchen an den Furcalästen. Das Männchen dreht sich herum, so dass die Köpfe in entgegengesetzte Richtungen weisen. Mit dem rechten 6. Thorakopoden wird das Abdomen des Weibchens ergriffen und mit dem linken 6. Thorakopoden eine Spermatophore an das Abdomen angeheftet. Bei der Eiablage entstehen Eisäcke, die dem Genitalsegment anhängen. Dort verbleiben die Eier bis zum Schlupf der Naupliuslarve.
Die Copepoda ernähren sich ausgesprochen vielseitig, entsprechend den einzelnen Lebensweisen. Alle Ernährungsweisen lassen sich jedoch von einer räuberischen ableiten. Es existieren etwa Filtrierer oder Arten, die auch einzelne, kleine Nahrungspartikel aktiv ergreifen können. Viele Arten beherrschen jedoch beides. Herbivore Arten können Algen herab zu einer Größe von 5 µm filtrieren. Carnivore Arten filtern Tiere von weniger als 30–50 µm, größere werden aktiv ergriffen. Bei Kommensalen und Parasiten reicht die Bandbreite vom einfachen Abfallverwerter bis zum stationären Blutsauger. Als Nahrung dient den Filtrierern Detritus und das gesamte Spektrum an Phytoplankton, wobei gewisse Arten, je nach ökologischen Bedingungen, ganz spezifisch die Nahrung auswählen. Die Räuber nehmen sämtliche Beute zu sich, die sie überwältigen können. Dies sind etwa gehäuselose Protozoa, andere Copepoda bis hin zu Fischbrut. Ecto- bzw. Endoparasiten ernähren sich von Hautfetzen und Schleim bzw. von der aufgenommenen Blutflüssigkeit.
Die Copepoda sind vermutlich die individuenreichste Gruppe der Metazoa auf der Erde. In den oberen Schichten fast aller Meeresoberflächen stellen sie 90 % des gesamten Zooplanktons. Da sie manchmal in dichten Schwärmen (etwa 10.000 Individuen pro m³ bei Pseudocalanus-Arten) auftreten, spielen sie für die Ernährung vieler wirtschaftlich genutzter Fische eine entscheidende Rolle. Auch im Süßwasser sind Copepoden die wichtigste Nahrungsquelle für Fische. Andererseits können carnivore Copepoda im Süßwasser auch an der Fischbrut Schaden anrichten. Die Nahrungskette ist sehr kurz, da viele Copepoda sich vom Phytoplankton ernähren und dann direkt von Fischen verzehrt werden. Tote Copepoda werden sehr schnell zersetzt. Bei 22 °C Wassertemperatur geschieht dies in drei Tagen.
Für das Taxon gelten folgende Autapomorphien
Die Copepoden stellen eine Unterklasse in der Klassifikation der Krebstiere (Crustacea) dar. Huys & Boxshall unterscheiden ursprünglich 10 Ordnungen, wobei die Einteilung in Taxa gleichen Ranges auf Ordnungsebene problematisch ist[2]. Die Einbeziehung der Ordnung Poecilostomatoida in die Cyclopoida, die 2004 vorgeschlagen worden war,[3] hat sich nicht durchgesetzt. Somit ergeben sich für die innere Systematik folgende 10 Ordnungen:
Nach Huys & Boxshall ergibt sich aus den Verwandtschaftsverhältnissen folgendes Kladogramm:
Ruderfußkrebse (Copepoda) N.N.Calanoida
Mormonilloida
Harpacticoida
Siphonostomatoida
Monstrilloida
Misophrioida
Cyclopoida
Gelyelloida
Platycopioida
Der Artikel beruht hauptsächlich auf folgenden Unterlagen:
Ruderfußkrebse (Copepoda) sind kleine Krebse (Crustacea), die marine und limnische Gewässer besiedeln. Sie bilden mit rund 13.000 Morphospezies die artenreichste Gruppe der Crustaceen und stellen den größten Anteil des marinen Zooplanktons dar. Sie sind daher ein sehr wichtiges Element der marinen Nahrungskette. Sie leben als Filtrierer, Räuber oder Parasiten. Bei Letzteren kann aufgrund der Lebensweise eine stark modifizierte Morphologie auftreten.
Copepoda ya iku kéwan saku anggota crustacea cilik kang lumrahé ditemoni ning laut lan asring uga ditemoni ning kabèh habitat banyu tawar lan ngrombol mbentuk sumber protéin paling akèh ning samudra.[1] Spesiesé sing paling akèh ya iku plankton, nanging akèh uga spésies benthos lan ana spésies kontinental bisa urip ning habitat limno-terestrial lan liyané ning laladan kang restrial teles, kaya ta rawa.
Copepoda ya iku kéwan saku anggota crustacea cilik kang lumrahé ditemoni ning laut lan asring uga ditemoni ning kabèh habitat banyu tawar lan ngrombol mbentuk sumber protéin paling akèh ning samudra. Spesiesé sing paling akèh ya iku plankton, nanging akèh uga spésies benthos lan ana spésies kontinental bisa urip ning habitat limno-terestrial lan liyané ning laladan kang restrial teles, kaya ta rawa.
Eshkakoyoqli qisqichbaqalar (Soreroba) — qisqichbaqasimonlar sinfiga mansub umurtqasiz hayvonlar turkumi (baʼzan kenja sinfi). Uz. 0,1 mm — 3 sm, ayrim parazit turlari 30 sm gacha. Gavdasi murakkab bosh (unga 1koʻkrak boʻgʻimi ham kiradi), 5 boʻgʻimli koʻkrak va 4 boʻgʻimli qoringa boʻlinadi. Qorin boʻlimida oyoqdari boʻlmaydi; bu boʻlim ayri oʻsimtali telson bilan tugaydi. Faqat nauplius koʻzchasi bor. Antennulalari uzun boʻlib, sezgi organi, suzish va suvda muallaq turish vazifasini bajaradi. E. suzganda koʻkrak oyoklar eshkak rolini oʻynaydi (nomi shundan). Tana yuzasi orqali nafas oladi. E. baliqlar va ularning chavoqlari, moʻylovli kitlar va boshqalar suv hayvonlari uchun oziq. Bir qancha turlari suv hayvonlari, koʻproq baliqlarda parazitlik qiladi. Tana shakli yashash tarziga bogʻliq. Suv qaʼrida yashaydigan Eshkakoyoqli qisqichbaqalar tanasi suyri shaklda boʻlib, juda uzun tukli oʻsimtalarga ega; suv tubida yashaydigan Eshkakoyoqli qisqichbaqalar tanasi yassi, tana oʻsimtalari kalta; parazit turlarining tanasi yashash tarzi taʼsirida regressiv oʻzgarib, kuchli morfofiziologik degeneratsiyaga uchraydi (sakkulina). Ayrim turlari odam organizmida parazitlik qiluvchi tasmasimon va toʻgarak chuvalchanglarning oraliq xoʻjayini (mas, chuchuk suvlarda yashaydigan sikloplar tanasida keng tasmasimon chuvalchang va rishta lichinkalari uchraydi). 6000 dan ortiq turi maʼlum. Chuchuk suv va dengizlarda planktonning asosiy qismini tashkil etadi. Chuchuv suv havzalarida, jumladan Oʻzbekistonda sikloplar, dengizlarda kalanuslar keng tarqalgan.
Τα κωπήποδα είναι μια ομάδα μικρών καρκινοειδών που βρίσκονται στη θάλασσα και σχεδόν σε όλους τους βιότοπους του γλυκού νερού και αποτελούν τη μεγαλύτερη πηγή πρωτεΐνης στους ωκεανούς [1]. Πολλά είδη είναι πλαγκτικά, αλλά τα περισσότερα είναι βενθικά. Υπάρχουν και μερικά ηπειρωτικά είδη που μπορούν να ζήσουν στους λιμναίους επίγειους βιότοπους και άλλα υγρά επίγεια οικοσυστήματα, όπως τα έλη, κάτω από τα πεσμένα φύλλα των υγρών δασών, βάλτους, ρυάκια, τις εφήμερες λίμνες και τις λακκούβες, υγρά βρύα, ή τις κοιλότητες των φυτών γεμάτες με νερό (φυτοτέλματα), όπως τα βρομελιοειδή (bromeliaceae) και τα φυτά-αμφορείς (Heliamphora). Πολλά ζουν υπόγεια σε θαλάσσιες σπηλιές και σπηλιές με γλυκά νερά, σε καταβόθρες, ή στο βυθό ρυακιών. Μερικά κωπήποδα είναι παρασιτικά και επικολλούνται σε ψάρια, καρχαρίες, θαλάσσια θηλαστικά, και πολλά είδη ασπόνδυλων όπως τα μαλάκια, χιτωνοφόρα, ή κοράλλια. Το κωπήποδα χρησιμοποιούνται μερικές φορές ως βιοενδείκτες.
Μπορεί να είναι φορείς του παρασιτικού σκώληκα Diphyllobothrium latum (Βοθριοκέφαλος ο πλατύς), που παρασιτεί στο δωδεκαδάκτυλο και προκαλεί έλλειψη βιταμίνης Β12.
Τα κωπήποδα είναι μια ομάδα μικρών καρκινοειδών που βρίσκονται στη θάλασσα και σχεδόν σε όλους τους βιότοπους του γλυκού νερού και αποτελούν τη μεγαλύτερη πηγή πρωτεΐνης στους ωκεανούς . Πολλά είδη είναι πλαγκτικά, αλλά τα περισσότερα είναι βενθικά. Υπάρχουν και μερικά ηπειρωτικά είδη που μπορούν να ζήσουν στους λιμναίους επίγειους βιότοπους και άλλα υγρά επίγεια οικοσυστήματα, όπως τα έλη, κάτω από τα πεσμένα φύλλα των υγρών δασών, βάλτους, ρυάκια, τις εφήμερες λίμνες και τις λακκούβες, υγρά βρύα, ή τις κοιλότητες των φυτών γεμάτες με νερό (φυτοτέλματα), όπως τα βρομελιοειδή (bromeliaceae) και τα φυτά-αμφορείς (Heliamphora). Πολλά ζουν υπόγεια σε θαλάσσιες σπηλιές και σπηλιές με γλυκά νερά, σε καταβόθρες, ή στο βυθό ρυακιών. Μερικά κωπήποδα είναι παρασιτικά και επικολλούνται σε ψάρια, καρχαρίες, θαλάσσια θηλαστικά, και πολλά είδη ασπόνδυλων όπως τα μαλάκια, χιτωνοφόρα, ή κοράλλια. Το κωπήποδα χρησιμοποιούνται μερικές φορές ως βιοενδείκτες.
Μπορεί να είναι φορείς του παρασιτικού σκώληκα Diphyllobothrium latum (Βοθριοκέφαλος ο πλατύς), που παρασιτεί στο δωδεκαδάκτυλο και προκαλεί έλλειψη βιταμίνης Β12.
Ишкәгаяклы́ кысласыманна́р (Copepoda) - кысласыманнар классыннан умырткасызлар отряды. 1800 гә якын төре билгеле, Татарстан территориясендә 60 чамасы. Буынлы, кабырчыксыз, башкүкрәк, баш һәм корсак өлешенә бүленгән гәүдәсе белән аерылып торучы вак кыслалар. 2 пар озын мыекчасы һәм кысла хәрәкәтләнгәндә ишкәк сыман (исеме шуннан) йөреш ясаучы 5 пар күкрәк очлыгы бар.
Гәүдәсе 0,5-3 мм озынлыкта, ана затлар аталарына караганда эрерәк. Ана ишкәгаяклы кысласыманнар аталанган йомыркаларын корсак астындагы махсус капчыкларда йөртә. Личинкалары метаморфоз юлы белән үсә һәм науплиус, копеподит стадияләрен уза. Өч асотрядка бүленәләр: каланоидлар (Calanoida), циклопоидлар (Cyclopoida) һәм гарпактикоидлар (Harpacticoida). Туклану характеры буенча беренчеләре - фильтрлаучыларга, икенчеләре - фильтрлаучы һәм ерткычларга, өченчеләре җыючы-детритофагларга карый. ишкәгаяклы кысласыманнар акмый торган һәм агымлы сулыклар зоопланктонының мөһим компонентлары, су төбе катламнарында да, су үсемлекләре арасында да яшәргә мөмкиннәр, алар - планктон белән тукланучы балыкларның яраткан азыгы. Циклопларның кайбер төрләре кеше паразитларының (мәсәлән, лентец, ришта) арадаш хуҗалары булып тора.
Ergasilus төренең циклоплары балыклар паразиты буларак билгеле һәм аларга зур зыян китерергә мөмкиннәр.

Ишкәгаяклы́ кысласыманна́р (Copepoda) - кысласыманнар классыннан умырткасызлар отряды. 1800 гә якын төре билгеле, Татарстан территориясендә 60 чамасы. Буынлы, кабырчыксыз, башкүкрәк, баш һәм корсак өлешенә бүленгән гәүдәсе белән аерылып торучы вак кыслалар. 2 пар озын мыекчасы һәм кысла хәрәкәтләнгәндә ишкәк сыман (исеме шуннан) йөреш ясаучы 5 пар күкрәк очлыгы бар.
Гәүдәсе 0,5-3 мм озынлыкта, ана затлар аталарына караганда эрерәк. Ана ишкәгаяклы кысласыманнар аталанган йомыркаларын корсак астындагы махсус капчыкларда йөртә. Личинкалары метаморфоз юлы белән үсә һәм науплиус, копеподит стадияләрен уза. Өч асотрядка бүленәләр: каланоидлар (Calanoida), циклопоидлар (Cyclopoida) һәм гарпактикоидлар (Harpacticoida). Туклану характеры буенча беренчеләре - фильтрлаучыларга, икенчеләре - фильтрлаучы һәм ерткычларга, өченчеләре җыючы-детритофагларга карый. ишкәгаяклы кысласыманнар акмый торган һәм агымлы сулыклар зоопланктонының мөһим компонентлары, су төбе катламнарында да, су үсемлекләре арасында да яшәргә мөмкиннәр, алар - планктон белән тукланучы балыкларның яраткан азыгы. Циклопларның кайбер төрләре кеше паразитларының (мәсәлән, лентец, ришта) арадаш хуҗалары булып тора.
Ergasilus төренең циклоплары балыклар паразиты буларак билгеле һәм аларга зур зыян китерергә мөмкиннәр.
Ишкәгаяҡтар (лат. Copepoda) — ҡыҫала һымаҡтар класынан умыртҡаһыҙҙар отряды; сөсө һәм тоҙло һыуҙарҙа йәшәй торған түбән ҡыҫалалар [1] . 1800 -гә яҡын төрө билдәле.
Быуынлы, ҡабырсаҡһыҙ, башкүкрәк, баш һәм ҡорһаҡ өлөшөнә бүленгән кәүҙәһе менән айырылып тороусы ваҡ ҡыҫалалар. 2 пар оҙон мыйыҡсаһы һәм ҡыҫала хәрәкәтләнгәндә ишкәк һымаҡ (исеме шунан) йөреш яһаусы 5 пар күкрәк ослоғо бар. Кәүҙәһе 0,5-3 мм оҙонлоҡта, инә заттар аталарына ҡарағанда эрерәк. Инә ишкәгаяҡлы ҡыҫала һымактар аталанған йомортҡаларын ҡорһаҡ аҫтындағы махсус ҡапсыҡтарҙа йөрөтә. Личинкалары метаморфоз юлы менән үҫә һәм науплиус, копеподит стадияләрын уҙа. Өч аҫ отрядҡа бүленәләр: каланоидтар (Calanoida), циклопоидтар (Cyclopoida) һәм гарпактикоидтар (Harpacticoida). Туҡланыу характеры буйынса беренселәре − фильтрлаусыларға, икенселәре − фильтрлаусы һәм йырткыстарға, өсөнсөләре йыйыусы −детритофагтарға ҡарай. Ишкәгаяҡлы ҡыҫала һымаҡтар аҡмай торған һәм ағымлы һыулыҡтар зоопланктонының мөһим компоненттары, һыу төбө ҡатламдарында ла, һыу үҫемлектәре араһында ла йәшәргә мөмкин, улар - планктон менән туҡланыусы балыҡтарҙың яратҡан аҙығы. Циклопларҙың ҡайһы бер төрҙәре кеше паразиттарының (мәҫәлән, лентец, ришта) аралаш хужалары булып тора. Ergasilus төрөнөң циклоптары балыҡтар паразиты булараҡ билдәле һәм уларга ҙур зыян килтерергә мөмкин.
Ишкәгаяҡтар (лат. Copepoda) — ҡыҫала һымаҡтар класынан умыртҡаһыҙҙар отряды; сөсө һәм тоҙло һыуҙарҙа йәшәй торған түбән ҡыҫалалар . 1800 -гә яҡын төрө билдәле.
Copepods (/ˈkoʊpɪpɒd/; meaning "oar-feet") are a group of small crustaceans found in nearly every freshwater and saltwater habitat. Some species are planktonic (inhabiting sea waters), some are benthic (living on the ocean floor), a number of species have parasitic phases, and some continental species may live in limnoterrestrial habitats and other wet terrestrial places, such as swamps, under leaf fall in wet forests, bogs, springs, ephemeral ponds, puddles, damp moss, or water-filled recesses of plants (phytotelmata) such as bromeliads and pitcher plants. Many live underground in marine and freshwater caves, sinkholes, or stream beds. Copepods are sometimes used as biodiversity indicators.
As with other crustaceans, copepods have a larval form. For copepods, the egg hatches into a nauplius form, with a head and a tail but no true thorax or abdomen. The larva molts several times until it resembles the adult and then, after more molts, achieves adult development. The nauplius form is so different from the adult form that it was once thought to be a separate species. The metamorphosis had, until 1832, led to copepods being misidentified as zoophytes or insects (albeit aquatic ones), or, for parasitic copepods, 'fish lice'.[1]
Copepods are currently assigned to the class Copepoda within the superclass Multicrustacea in the subphylum Crustacea.[2] An alternative treatment is as a subclass belonging to class Hexanauplia.[3] They are divided into 10 orders. Some 13,000 species of copepods are known, and 2,800 of them live in fresh water.[4]

Copepods vary considerably, but are typically 1 to 2 mm (1⁄32 to 3⁄32 in) long, with a teardrop-shaped body and large antennae. Like other crustaceans, they have an armoured exoskeleton, but they are so small that in most species, this thin armour and the entire body is almost totally transparent. Some polar copepods reach 1 cm (1⁄2 in). Most copepods have a single median compound eye, usually bright red and in the centre of the transparent head. Subterranean species may be eyeless, and members of the genera Copilia and Corycaeus possess two eyes, each of which has a large anterior cuticular lens paired with a posterior internal lens to form a telescope.[5][6][7] Like other crustaceans, copepods possess two pairs of antennae; the first pair is often long and conspicuous.
Free-living copepods of the orders Calanoida, Cyclopoida, and Harpacticoida typically have a short, cylindrical body, with a rounded or beaked head, although considerable variation exists in this pattern. The head is fused with the first one or two thoracic segments, while the remainder of the thorax has three to five segments, each with limbs. The first pair of thoracic appendages is modified to form maxillipeds, which assist in feeding. The abdomen is typically narrower than the thorax, and contains five segments without any appendages, except for some tail-like "rami" at the tip.[8] Parasitic copepods (the other seven orders) vary widely in morphology and no generalizations are possible.
Because of their small size, copepods have no need of any heart or circulatory system (the members of the order Calanoida have a heart, but no blood vessels), and most also lack gills. Instead, they absorb oxygen directly into their bodies. Their excretory system consists of maxillary glands.
The second pair of cephalic appendages in free-living copepods is usually the main time-averaged source of propulsion, beating like oars to pull the animal through the water. However, different groups have different modes of feeding and locomotion, ranging from almost immotile for several minutes (e.g. some harpacticoid copepods) to intermittent motion (e.g., some cyclopoid copepods) and continuous displacements with some escape reactions (e.g. most calanoid copepods).

Some copepods have extremely fast escape responses when a predator is sensed, and can jump with high speed over a few millimetres. Many species have neurons surrounded by myelin (for increased conduction speed), which is very rare among invertebrates (other examples are some annelids and malacostracan crustaceans like palaemonid shrimp and penaeids). Even rarer, the myelin is highly organized, resembling the well-organized wrapping found in vertebrates (Gnathostomata). Despite their fast escape response, copepods are successfully hunted by slow-swimming seahorses, which approach their prey so gradually, it senses no turbulence, then suck the copepod into their snout too suddenly for the copepod to escape.[9]
Finding a mate in the three-dimensional space of open water is challenging. Some copepod females solve the problem by emitting pheromones, which leave a trail in the water that the male can follow.[10] Copepods experience a low Reynolds number and therefore a high relative viscosity. One foraging strategy involves chemical detection of sinking marine snow aggregates and taking advantage of nearby low-pressure gradients to swim quickly towards food sources.[11]
Most free-living copepods feed directly on phytoplankton, catching cells individually. A single copepod can consume up to 373,000 phytoplankton per day.[12] They generally have to clear the equivalent to about a million times their own body volume of water every day to cover their nutritional needs.[13] Some of the larger species are predators of their smaller relatives. Many benthic copepods eat organic detritus or the bacteria that grow in it, and their mouth parts are adapted for scraping and biting. Herbivorous copepods, particularly those in rich, cold seas, store up energy from their food as oil droplets while they feed in the spring and summer on plankton blooms. These droplets may take up over half of the volume of their bodies in polar species. Many copepods (e.g., fish lice like the Siphonostomatoida) are parasites, and feed on their host organisms. In fact, three of the 10 known orders of copepods are wholly or largely parasitic, with another three comprising most of the free-living species.[14]
Most nonparasitic copepods are holoplanktonic, meaning they stay planktonic for all of their lifecycles, although harpacticoids, although free-living, tend to be benthic rather than planktonic. During mating, the male copepod grips the female with his first pair of antennae, which is sometimes modified for this purpose. The male then produces an adhesive package of sperm and transfers it to the female's genital opening with his thoracic limbs. Eggs are sometimes laid directly into the water, but many species enclose them within a sac attached to the female's body until they hatch. In some pond-dwelling species, the eggs have a tough shell and can lie dormant for extended periods if the pond dries up.[8]
Eggs hatch into nauplius larvae, which consist of a head with a small tail, but no thorax or true abdomen. The nauplius moults five or six times, before emerging as a "copepodid larva". This stage resembles the adult, but has a simple, unsegmented abdomen and only three pairs of thoracic limbs. After a further five moults, the copepod takes on the adult form. The entire process from hatching to adulthood can take a week to a year, depending on the species and environmental conditions such as temperature and nutrition (e.g., egg-to-adult time in the calanoid Parvocalanus crassirostris is ~7 days at 25 °C (77 °F) but 19 days at 15 °C (59 °F).[15]
Copepods jump out of the water - porpoising. The biophysics of this motion has been described by Waggett and Buskey 2007 and Kim et al 2015.[16]
Planktonic copepods are important to global ecology and the carbon cycle. They are usually the dominant members of the zooplankton, and are major food organisms for small fish such as the dragonet, banded killifish, Alaska pollock, and other crustaceans such as krill in the ocean and in fresh water. Some scientists say they form the largest animal biomass on earth.[18] Copepods compete for this title with Antarctic krill (Euphausia superba). C. glacialis inhabits the edge of the Arctic icepack, especially in polynyas where light (and photosynthesis) is present, in which they alone comprise up to 80% of zooplankton biomass. They bloom as the ice recedes each spring. The ongoing large reduction in the annual ice pack minimum may force them to compete in the open ocean with the much less nourishing C. finmarchicus, which is spreading from the North Sea and the Norwegian Sea into the Barents Sea.[19]

Because of their smaller size and relatively faster growth rates, and because they are more evenly distributed throughout more of the world's oceans, copepods almost certainly contribute far more to the secondary productivity of the world's oceans, and to the global ocean carbon sink than krill, and perhaps more than all other groups of organisms together. The surface layers of the oceans are currently believed to be the world's largest carbon sink, absorbing about 2 billion tons of carbon a year, the equivalent to perhaps a third of human carbon emissions, thus reducing their impact. Many planktonic copepods feed near the surface at night, then sink (by changing oils into more dense fats)[20][21] into deeper water during the day to avoid visual predators. Their moulted exoskeletons, faecal pellets, and respiration at depth all bring carbon to the deep sea.
About half of the estimated 14,000 described species of copepods are parasitic[22] [23] and many have adapted extremely modified bodies for their parasitic lifestyles.[24] They attach themselves to bony fish, sharks, marine mammals, and many kinds of invertebrates such as corals, other crustaceans, molluscs, sponges, and tunicates. They also live as ectoparasites on some freshwater fish.[25]
In addition to being parasites themselves, copepods are subject to parasitic infection. The most common parasites are marine dinoflagellates of the genus Blastodinium, which are gut parasites of many copepod species.[26][27] Currently, 12 species of Blastodinium are described, the majority of which were discovered in the Mediterranean Sea.[26] Most Blastodinium species infect several different hosts, but species-specific infection of copepods does occur. Generally, adult copepod females and juveniles are infected.
During the naupliar stage, the copepod host ingests the unicellular dinospore of the parasite. The dinospore is not digested and continues to grow inside the intestinal lumen of the copepod. Eventually, the parasite divides into a multicellular arrangement called a trophont.[28] This trophont is considered parasitic, contains thousands of cells, and can be several hundred micrometers in length.[27] The trophont is greenish to brownish in color as a result of well-defined chloroplasts. At maturity, the trophont ruptures and Blastodinium spp. are released from the copepod anus as free dinospore cells. Not much is known about the dinospore stage of Blastodinium and its ability to persist outside of the copepod host in relatively high abundances.[29]
The copepod Calanus finmarchicus, which dominates the northeastern Atlantic coast, has been shown to be greatly infected by this parasite. A 2014 study in this region found up to 58% of collected C. finmarchicus females to be infected.[28] In this study, Blastodinium-infected females had no measurable feeding rate over a 24-hour period. This is compared to uninfected females which, on average, ate 2.93 × 104 cells per day.[28] Blastodinium-infected females of C. finmarchicus exhibited characteristic signs of starvation, including decreased respiration, fecundity, and fecal pellet production. Though photosynthetic, Blastodinium spp. procure most of their energy from organic material in the copepod gut, thus contributing to host starvation.[27] Underdeveloped or disintegrated ovaries and decreased fecal pellet size are a direct result of starvation in female copepods.[30] Parasitic infection by Blastodinium spp. could have serious ramifications on the success of copepod species and the function of entire marine ecosystems. Blastodinium parasitism is not lethal, but has negative impacts on copepod physiology, which in turn may alter marine biogeochemical cycles.
Freshwater copepods of the Cyclops genus are the intermediate host of the Guinea worm (Dracunculus medinensis), the nematode that causes dracunculiasis disease in humans. This disease may be close to being eradicated through efforts by the U.S. Centers for Disease Control and Prevention and the World Health Organization.[31]
Despite their modern abundance, due to their small size and fragility, copepods are extremely rare in the fossil record. The oldest known fossils of copepods are from the late Carboniferous (Pennsylvanian) of Oman, around 303 million years old, which were found in a clast of bitumen from a glacial diamictite. The copepods present in the bitumen clast were likely residents of a subglacial lake which the bitumen had seeped upwards through while still liquid, before the clast subsequently soldified and was deposited by glaciers. Though most of the remains were undiagnostic, at least some likely belonged to the extant harpacticoid family Canthocamptidae, suggesting that copepods had already substantially diversified by this time.[32] Possible microfossils of copepods are known from the Cambrian of North America.[33][34] Transitions to parasitism have occurred within copepods independently at least 14 different times, with the oldest record of this being from damage to fossil echinoids done by cyclopoids from the Middle Jurassic of France, around 168 million years old.[35]
Live copepods are used in the saltwater aquarium hobby as a food source and are generally considered beneficial in most reef tanks. They are scavengers and also may feed on algae, including coralline algae. Live copepods are popular among hobbyists who are attempting to keep particularly difficult species such as the mandarin dragonet or scooter blenny. They are also popular to hobbyists who want to breed marine species in captivity. In a saltwater aquarium, copepods are typically stocked in the refugium.
Copepods are sometimes found in public main water supplies, especially systems where the water is not mechanically filtered,[36] such as New York City, Boston, and San Francisco.[37] This is not usually a problem in treated water supplies. In some tropical countries, such as Peru and Bangladesh, a correlation has been found between copepods' presence and cholera in untreated water, because the cholera bacteria attach to the surfaces of planktonic animals. The larvae of the guinea worm must develop within a copepod's digestive tract before being transmitted to humans. The risk of infection with these diseases can be reduced by filtering out the copepods (and other matter), for example with a cloth filter.[38]
Copepods have been used successfully in Vietnam to control disease-bearing mosquitoes such as Aedes aegypti that transmit dengue fever and other human parasitic diseases.[39][40]
The copepods can be added to water-storage containers where the mosquitoes breed.[36] Copepods, primarily of the genera Mesocyclops and Macrocyclops (such as Macrocyclops albidus), can survive for periods of months in the containers, if the containers are not completely drained by their users. They attack, kill, and eat the younger first- and second-instar larvae of the mosquitoes. This biological control method is complemented by community trash removal and recycling to eliminate other possible mosquito-breeding sites. Because the water in these containers is drawn from uncontaminated sources such as rainfall, the risk of contamination by cholera bacteria is small, and in fact no cases of cholera have been linked to copepods introduced into water-storage containers. Trials using copepods to control container-breeding mosquitoes are underway in several other countries, including Thailand and the southern United States. The method, though, would be very ill-advised in areas where the guinea worm is endemic.
The presence of copepods in the New York City water supply system has caused problems for some Jewish people who observe kashrut. Copepods, being crustaceans, are not kosher, nor are they quite small enough to be ignored as nonfood microscopic organisms, since some specimens can be seen with the naked eye. When a group of rabbis in Brooklyn, New York, discovered the copepods in the summer of 2004, they triggered such debate in rabbinic circles that some observant Jews felt compelled to buy and install filters for their water.[41] The water was ruled kosher by posek Yisrael Belsky.[42]
The Nickelodeon television series SpongeBob SquarePants features a copepod named Sheldon J. Plankton as a recurring character.[43]
Copepods (/ˈkoʊpɪpɒd/; meaning "oar-feet") are a group of small crustaceans found in nearly every freshwater and saltwater habitat. Some species are planktonic (inhabiting sea waters), some are benthic (living on the ocean floor), a number of species have parasitic phases, and some continental species may live in limnoterrestrial habitats and other wet terrestrial places, such as swamps, under leaf fall in wet forests, bogs, springs, ephemeral ponds, puddles, damp moss, or water-filled recesses of plants (phytotelmata) such as bromeliads and pitcher plants. Many live underground in marine and freshwater caves, sinkholes, or stream beds. Copepods are sometimes used as biodiversity indicators.
As with other crustaceans, copepods have a larval form. For copepods, the egg hatches into a nauplius form, with a head and a tail but no true thorax or abdomen. The larva molts several times until it resembles the adult and then, after more molts, achieves adult development. The nauplius form is so different from the adult form that it was once thought to be a separate species. The metamorphosis had, until 1832, led to copepods being misidentified as zoophytes or insects (albeit aquatic ones), or, for parasitic copepods, 'fish lice'.
Kopepodoj (signife "remil-piedoj") estas grupo de malgrandaj krustuloj troviĝantaj en maroj kaj apudaj habitatoj de nesala akvo. Kelkaj specioj estas el planktono (flosantaj en maraj akvoj), kelkaj estas bentosaj (loĝantaj sur la oceana grundo), kaj kelkaj kontinentaj specioj povas loĝi en ŝlim-teraj habitatoj kaj aliaj en malsekaj teraj lokoj, kiaj marĉoj, sub folia subkreskaĵaro en humidaj arbaroj, fontoj, porptempaj lagetoj kaj flokoj, marĉa musko, aŭ akvujoj (fitotelmatoj) de plantoj kiaj la bromeliacoj kaj aliaj karnovoraj plantoj. Multaj vivas subgrunde en maraj kaj nemaraj kavoj, akvotruoj, aŭ rojoj. Kopepodoj estas foje uzataj kiel bioindikiloj.
Kopepodoj formas subklason apartenantan al la subfilumo de krustuloj. Kopepodoj estas dividataj en dek ordoj. Oni konas ĉirkaŭ 13,000 speciojn de kopepodoj, kaj 2,800 el ili loĝas en nesalaj akvoj.[1]
Kopepodoj (signife "remil-piedoj") estas grupo de malgrandaj krustuloj troviĝantaj en maroj kaj apudaj habitatoj de nesala akvo. Kelkaj specioj estas el planktono (flosantaj en maraj akvoj), kelkaj estas bentosaj (loĝantaj sur la oceana grundo), kaj kelkaj kontinentaj specioj povas loĝi en ŝlim-teraj habitatoj kaj aliaj en malsekaj teraj lokoj, kiaj marĉoj, sub folia subkreskaĵaro en humidaj arbaroj, fontoj, porptempaj lagetoj kaj flokoj, marĉa musko, aŭ akvujoj (fitotelmatoj) de plantoj kiaj la bromeliacoj kaj aliaj karnovoraj plantoj. Multaj vivas subgrunde en maraj kaj nemaraj kavoj, akvotruoj, aŭ rojoj. Kopepodoj estas foje uzataj kiel bioindikiloj.
Los copépodos (Copepoda, del griego «cope», remo, y «poda», pata) son una subclase de crustáceos maxilópodos de pequeño tamaño, muy extendidos por todo el planeta, principalmente formando parte del zooplancton, aunque también podemos encontrar especies semiterrestres. Existen especies adaptadas a casi todo tipo de ambientes acuáticos (dulce, salobre, salada, aguas antárticas, etc.). Además de haber podido colonizar una innumerable cantidad de nichos en todo el mundo, son considerados los artrópodos más abundantes.
Se conocen unas 12 000 especies, la mayoría de ellas son de vida libre, pero algunos géneros se han adaptado a estilos de vida comensal e incluso al parasitismo, utilizando un enorme rango de hospedadores para el desarrollo de su ciclo vital, a veces complejo. Concretamente, estos mesoparásitos (ya que suelen estar insertos parcialmente en los tejidos del huésped) desarrollan ciclos de vida con hospedadores secundarios y terciarios afectando a cetáceos, peces y moluscos principalmente. Este aspecto los convierten en importantes partícipes de las complejas redes tróficas que se forman entre los huéspedes, siendo relativamente fácil la transmisión y dispersión del parásito. Los copépodos per se son considerados a veces un problema para la acuicultura, ya que la alta densidad de hospedadores facilita una elevada concentración de copépodos.
Hay interacciones muy curiosas, un ejemplo es el cestodo Triaenophorus crassus, que es capaz de provocar cambios de comportamiento en su hospedador intermediario del género[1] que lo llevan a ocupar nichos que no suelen frecuentar y con ello se facilita su ingestión por especies concretas de peces. Actualmente, se ha propuesto el uso de algunas especies de copépodos para el control biológico.
la gran mayoría nada libremente; sin embargo, algunas especies se han convertido en parásitas. Muchas de estas últimas conservan un aspecto similar al de las formas libres, al tiempo que otras han llegado al extremo de perder sus características de crustáceo e incluso de artrópodo, notándose mayormente esta situación en el caso de las hembras. Para poder clasificar estas últimas especies dentro de los copépodos, se parte del estudio de machos jóvenes, los cuales no están muy modificados.
Los copépodos carecen de caparazón, y su desarrollo se inicia a partir de una larva nauplio. Dos géneros copépodos de agua dulce muy estudiados son Cyclops y Diaptomus, ambos ofrecen un cuadro general del grupo bastante amplio. En el caso de las especies del género Cyclops, su aspecto es el de un crustáceo decápodo, con una porción anterior con forma de pera que incluye la región cefálica y los dos primeros segmentos del tórax. A continuación se encuentran tres segmentos del tórax, móviles, y por último el abdomen, sin apéndices y que termina en una cola ahorquillada con filamentos plumosos. Tienen un solo ojo, que evolucionó de dos ojos que se fusionaron, y las antenas principales, muy articuladas son mucho mayores que las secundarias. Poseen 4 pares de apéndices torácicos que les sirven para nadar y vestigios de un quinto par que en otras especies de copépodos son más notorios. Adicionalmente a las mandíbulas y maxilas, el primer par de apéndices torácicos ha evolucionado hasta convertirse en maxilas secundarias.
La gran mayoría de los copépodos son de cuerpo transparente, aun cuando los hay con cierta coloración como los Anomalocera. Como hecho curioso, se puede notar que algunas especies que son traslúcidas, llegan a colorear abundantemente el agua debido a la difracción de la luz al pasar por sus cuerpos. Asimismo algunas especies marinas son luminosas por sí mismas por poseer en sus cuerpos alguna sustancia fosforescente.
Los copépodos son tan abundantes en el agua de mar que cualquier muestra de plancton tiene siempre ejemplares de este grupo. Están distribuidos a lo largo de todas las latitudes, incluyendo las antárticas. Muchas de las especies cuentan con apéndices plumosos que en muchos casos pueden llegar a superar el tamaño del animal hasta por 4 o 6 veces. En agua dulce también están ampliamente distribuidos, en forma tan amplia como los cladóceros.
Dentro de los copépodos hay una gran cantidad de especies parásitas, más que en ningún otro grupo de crustáceos. Son muy conocidos los llamados piojos de los peces de la familia de los calígidos, que se caracterizan por el cuerpo ancho y deprimido que les facilita adherirse a su huésped fácilmente. Una particularidad de los calígidos es que en sus primeras etapas de desarrollo nadan libremente, y solo inician su etapa parasitaria cuando son adultos jóvenes. Permanecen parásitos sin facultades natatorias durante un período de tiempo determinado por etapas de crecimiento y apareamiento y por último retornan a la vida libre, recuperando sus capacidades natatorias.
Otro parásito de costumbres curiosas es el Lernacea, cuyas larvas que nacen en forma de nauplio e inician su vida parásita sobre un pez de la familia de los pleuronéctidos, como el lenguado; sus apéndices se reducen hasta quedar convertidos en meros muñones. Cuando llega la época de apareamiento, el animal se desprende del huésped. Después de aparearse los machos mueren, y las hembras buscan un nuevo huésped, generalmente un bacalao o una merluza, se fijan en las branquias e inician un proceso de degeneración tal, que llegan a perder totalmente los apéndices hasta quedar convertidas en sacos sin ninguna apariencia de crustáceos.
Copépodos como transmisores de enfermedades.
Debido a su gran número, diversidad y diferentes implicaciones en procesos ecológicos son organismos que directa o indirectamente se cruzan con el ser humano. De hecho algunas especies (mayoritariamente de agua dulce) representan vías de transmisión directas o indirectas de parásitos oportunistas que pueden infectar a humanos y a otros vertebrados, convirtiéndose en vectores de algunos de estos parásitos (nematodes y plathihelmintos sobre todo). Las principales vías de ingesta de copépodos en el hombre son la ingesta de agua (su impresionante número hace muy rara la ausencia de copépodos en muestras de ríos y otras fuentes de agua dulce naturales), o la de otros organismos que ya los hayan consumido (pescado y otros huéspedes que transmiten el parásito al comerse crudos o poco cocinados, etc.). Estas vías permiten el acceso de platelmintos y nematodes al ser humano y le puede provocar a este serios problemas e incluso la muerte. Por ello, la profilaxis es de vital importancia para acabar con los casos puntuales de parasitosis, y para comenzar a elaborar proyectos encaminados a sistemas de filtración y depuración de agua, así como promover el consumo adecuado de los animales que podrían transmitir el parásito al ser humano o a sus animales de compañía.
Parásitos del ser humano que emplean copépodos como hospedadores intermediarios.
Plathihelmintos
Nematodos
Bacterias
Actualmente, el estudio de las dinámicas poblacionales del fitoplancton y zooplancton vía satélite (detectando cambios en la cantidad de clorofila en el agua) en zonas costeras como el golfo de Bengala permite prevenir posibles nuevas epidemias, ya que las bacterias se asocian a estos organismos.[6]
Según Martin & Davis (2001),[7] los copépodos incluyen los siguientes grupos:
Infraclase Progymnoplea Lang, 1948
Infraclase Neocopepoda Huys & Boxshall, 1991
Los copépodos (Copepoda, del griego «cope», remo, y «poda», pata) son una subclase de crustáceos maxilópodos de pequeño tamaño, muy extendidos por todo el planeta, principalmente formando parte del zooplancton, aunque también podemos encontrar especies semiterrestres. Existen especies adaptadas a casi todo tipo de ambientes acuáticos (dulce, salobre, salada, aguas antárticas, etc.). Además de haber podido colonizar una innumerable cantidad de nichos en todo el mundo, son considerados los artrópodos más abundantes.
Se conocen unas 12 000 especies, la mayoría de ellas son de vida libre, pero algunos géneros se han adaptado a estilos de vida comensal e incluso al parasitismo, utilizando un enorme rango de hospedadores para el desarrollo de su ciclo vital, a veces complejo. Concretamente, estos mesoparásitos (ya que suelen estar insertos parcialmente en los tejidos del huésped) desarrollan ciclos de vida con hospedadores secundarios y terciarios afectando a cetáceos, peces y moluscos principalmente. Este aspecto los convierten en importantes partícipes de las complejas redes tróficas que se forman entre los huéspedes, siendo relativamente fácil la transmisión y dispersión del parásito. Los copépodos per se son considerados a veces un problema para la acuicultura, ya que la alta densidad de hospedadores facilita una elevada concentración de copépodos.
Hay interacciones muy curiosas, un ejemplo es el cestodo Triaenophorus crassus, que es capaz de provocar cambios de comportamiento en su hospedador intermediario del género que lo llevan a ocupar nichos que no suelen frecuentar y con ello se facilita su ingestión por especies concretas de peces. Actualmente, se ha propuesto el uso de algunas especies de copépodos para el control biológico.
Aerjalalised ehk kopepoodid (Copepoda) on lülijalgsete hõimkonda kuuluva aerjalgsete klassi alamklass (võidakse vaadelda ka seltsina aerjalgsete alamklassis[viide?]).
Aerjalalisi leidub meres ja enamikus mageveeveekogudes. Paljud aerjalalised on planktilise eluviisiga, osa on bentilised. Esineb ka parasiitseid liike.
Planktilised aerjalalised on zooplanktoni oluline rühm, nad on toiduks kaladele, vaaladele, merelindudele ning teistele lülijalgsetele. Aerjalalised on tähtsad globaalökoloogias ja süsinikuringes. Mõnede allikate kohaselt on aerjalaliste biomass kõige suurem loomne biomass[viide?].
Aerjalalised on mikroskoopilised, nende pikkus on alla 1 mm kuni 5 mm[viide?]. Kehaehituses eristub pea, rindmik ja tagakeha. Pea on kokku kasvanud rindmiku esimese segmendiga. Pea eesotsas asub allapoole kaldus nokis ehk rostrum.
Tavaliselt on aerjalaliselt pea eesosas paaritu liitsilm ja lihtsilmana talitlev naupliussilm.
Nende eestundlad (antennid) on suhteliselt pikad ja osalevad ujumisel ja hõljumisel. Aerjalaliste liikumine pole sujuv, vaid tõukeline.
Enamik aerjalalised toituvad vee filtreerimisest (filtrijad), vähem on parasiite.
Aerjalalised on lahksugulised. Munadest kooruvat vastset nimetatakse vähikvastseks ehk naupliuseks. Elu jooksul kestuvad nad mitu korda.
Aerjalaliste alamklassi kuuluvad seltsid:
Aerjalalised ehk kopepoodid (Copepoda) on lülijalgsete hõimkonda kuuluva aerjalgsete klassi alamklass (võidakse vaadelda ka seltsina aerjalgsete alamklassis[viide?]).
Aerjalalisi leidub meres ja enamikus mageveeveekogudes. Paljud aerjalalised on planktilise eluviisiga, osa on bentilised. Esineb ka parasiitseid liike.
Planktilised aerjalalised on zooplanktoni oluline rühm, nad on toiduks kaladele, vaaladele, merelindudele ning teistele lülijalgsetele. Aerjalalised on tähtsad globaalökoloogias ja süsinikuringes. Mõnede allikate kohaselt on aerjalaliste biomass kõige suurem loomne biomass[viide?].
Kopepodoak (izen zientifikoa: Copepoda) itsasoan eta ur gezako ia edozein habitatetan bizi den krustazeo txikien talde bat dira. Espezie batzuk planktonikoak dira (itsasoko uretan igeri dabiltza), beste batzuk bentonikoak dira (ozeanoko hondoetan finko bizi dira) eta beste batzuk limno-lurtarrak dira, leku hezeetan biziz: padurak, hosto erorien azpian, ur-iturrietan, urtaroetan bakarrik sortzen diren putzu eta urmaeletan, goroldioen azpian, edo urez betetzen diren hainbat landareen artean. Lurpean ere bizi daitezke, kobazulo, sima edo antzekoetan. Kopepodoen barruan dauden animalia batzuk bioindikadoreak dira.
Hankajalkaiset (Copepoda) ovat pieniä äyriäisiä, joista suurin osa kuuluu planktoniin. Suurimmat hankajalkaiset voi nähdä paljain silmin, pääosa lajeista näkyy vain mikroskoopilla. Hankajalkaisilla on elinkaarensa aikana monta muodonmuutosta, ja jokaisen aikana ne vaihtavat kuorensa.[1] Plankton-hankajalkaiset ovat monien kalojen tärkeää ravintoa, muun muassa silakan.
Hankajalkaiset jaetaan kymmeneen lahkoon. Niitä tunnetaan noin 13 000 lajia, joista 2 800 elää makeassa vedessä, muut merissä.[2] Planktoniin kuuluvien lajien ohella hankajalkaisiin kuuluu pohjalla eläviä lajeja ja paljon muiden eläinten loisina eläviä lajeja.
Hankajalkaiset (Copepoda) ovat pieniä äyriäisiä, joista suurin osa kuuluu planktoniin. Suurimmat hankajalkaiset voi nähdä paljain silmin, pääosa lajeista näkyy vain mikroskoopilla. Hankajalkaisilla on elinkaarensa aikana monta muodonmuutosta, ja jokaisen aikana ne vaihtavat kuorensa. Plankton-hankajalkaiset ovat monien kalojen tärkeää ravintoa, muun muassa silakan.
Hankajalkaiset jaetaan kymmeneen lahkoon. Niitä tunnetaan noin 13 000 lajia, joista 2 800 elää makeassa vedessä, muut merissä. Planktoniin kuuluvien lajien ohella hankajalkaisiin kuuluu pohjalla eläviä lajeja ja paljon muiden eläinten loisina eläviä lajeja.
Les copépodes sont un groupe de petits crustacés, libres et parasites (externe ou interne d'organismes variés), vivant dans l'eau de mer et dans presque tous les habitats d'eau douce (lacs, marais, rivières, eaux souterraines). En mer, ils forment la base du plancton et la nourriture des poissons. D'autres espèces sont benthiques, ou encore parasites.
L'ordre des Calanoida domine dans le plancton marin, avec des genres tels que Copilia, Oncea, Porcellidium. On dénombre dans certaines mers plusieurs dizaines de milliers d'individus de Calanus finmarchicus par mètre carré de surface ; cette espèce domine le plancton de l'Atlantique Nord. En eau douce, c'est l'ordre des Cyclopoidas qui abonde. Quelques espèces sont adaptées aux eaux saumâtres.
Certains copépodes sont les espèces hôtes naturelles du vibrion du choléra (Vibrio cholerae) et donc vectrices du choléra, ce qui explique un lien entre changement climatique, épisodes climatiques chauds et épidémies de choléra[1] et le fait que divers facteurs environnementaux puissent agir écoépidémiologiquement sur le choléra dont les vibrions peuvent proliférer sur la surface d'algues où se nourrissent les copépodes, et in fine sur les cuticules de crustacés, copépodes notamment quand il y a pullulation d'algues vertes filamenteuses (également en contact avec des insectes à marée basse)[2]. Les changements environnementaux source de blooms planctoniques et proliférations bactériennes sont identifiés depuis quelques années et on a montré en 2007 que les vibrions pouvaient survivre dans des biofilms tridimensionnels dans l'environnement marin entre deux épidémies ou pullulations[2].
Les copépodes sont de petite taille. Ils n'ont ni branchies ni carapace, et ne présentent qu'un seul œil, médian, appelé œil nauplien. Ils nagent par petits sauts, à l'aide d'antennes natatoires. Les femelles sont reconnaissables à leurs deux sacs ovigènes latéraux. La larve sortant de l'œuf est appelée nauplius.
Certaines espèces de copépodes sont parasites de cnidaires, mollusques, crustacés, ascidies, poissons (pou du poisson), cétacés. La morphologie des espèces parasites est très différente de celle des espèces libres. Là où ils sont nombreux, ils peuvent constituer jusqu'à 60 % de la biomasse du zooplancton océanique[3].
Avec quelques exceptions pour les espèces parasites, leurs caractéristiques sont les suivantes :
Les copépodes planctoniques jouent un rôle important dans les réseaux trophiques marins et le cycle du carbone. Ils régulent les populations de protistes et d'algues unicellulaires. Ils contribuent au micromélange des couches d'eau par le battement incessant de leurs appendices natatoires.
Milne Edwards a nommé les copépodes (du grec ancien κώπη, kỗpê, «rame», et πούς, pous {au génitif ποδός, podos}, «pied») en 1830 en raison de la pièce squelettique dite copula qui relie la base des deux éléments de chaque paire des pattes thoraciques natatoires. C'est elle qui permet le battement synchrone de ces pattes [3].
Parmi environ 10 000 espèces connues, on distingue selon World Register of Marine Species (7 mars 2017)[4] :
De nombreux copépodes sont parasites d'espèces appartenant à presque tous les groupes animaux marins (des petits spongiaires aux plus grands cétacés). Parfois leur cycle parasitaire passe par plusieurs hôtes, par exemple chez les Pennellidae (en). Les modes parasitaires sont :
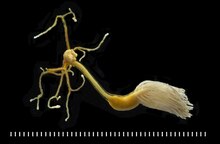
Lernaeenicus sprattae (en) parasitant un sprat (Sprattus sprattus).
Adulte de Lernaeolophus sultanus (Pennellidae), parasite du poisson Pristipomoides filamentosus.
Échelle : 1 mm par barre[6].
Les copépodes sont un groupe de petits crustacés, libres et parasites (externe ou interne d'organismes variés), vivant dans l'eau de mer et dans presque tous les habitats d'eau douce (lacs, marais, rivières, eaux souterraines). En mer, ils forment la base du plancton et la nourriture des poissons. D'autres espèces sont benthiques, ou encore parasites.
L'ordre des Calanoida domine dans le plancton marin, avec des genres tels que Copilia, Oncea, Porcellidium. On dénombre dans certaines mers plusieurs dizaines de milliers d'individus de Calanus finmarchicus par mètre carré de surface ; cette espèce domine le plancton de l'Atlantique Nord. En eau douce, c'est l'ordre des Cyclopoidas qui abonde. Quelques espèces sont adaptées aux eaux saumâtres.
Certains copépodes sont les espèces hôtes naturelles du vibrion du choléra (Vibrio cholerae) et donc vectrices du choléra, ce qui explique un lien entre changement climatique, épisodes climatiques chauds et épidémies de choléra et le fait que divers facteurs environnementaux puissent agir écoépidémiologiquement sur le choléra dont les vibrions peuvent proliférer sur la surface d'algues où se nourrissent les copépodes, et in fine sur les cuticules de crustacés, copépodes notamment quand il y a pullulation d'algues vertes filamenteuses (également en contact avec des insectes à marée basse). Les changements environnementaux source de blooms planctoniques et proliférations bactériennes sont identifiés depuis quelques années et on a montré en 2007 que les vibrions pouvaient survivre dans des biofilms tridimensionnels dans l'environnement marin entre deux épidémies ou pullulations.
Artrapód bídeach uisciúil. Tá foirmeacha a mhaireann go saor an-fhlúirseach san iliomad gnáthóg muirí is fionnuisce, ag déanamh naisc bhunúsaigh sa slabhra bia trí mhionphlandaí planctóin a ithe. Timpeall 9,000 speiceas sa bhfo-rang, cuid mhaith díobh comhthíosach le grúpaí eile ainmhithe nó seadánach orthu.
Os copépodos son un numerosísimo grupo de pequenos crustáceos, normalmente de menos de 1 mm de lonxitude, pero que representan ó redor do 80% do zooplancto mariño.
A maioría dos copépodos son mariños, pero hai moitas especies dulciacuícolas e unhas poucas que viven entre a vexetación húmida e nas películas de auga na terra. Tamén hai moitas especies parasitas de diferentes animais mariños e dulciacuícolas, especialmente de peixes.
Dentro da enorme variedade morfolóxica lóxica nun grupo zoolóxico tan amplo (hai descritas máis de 10.000 especies), presentan un corpo alongado, extremo caudal bifurcado, cinco pares de patas torácicas e carecen do coiraza dura propia dos crustáceos máis coñecidos.
O tronco está composto polo tórax e o abdome. O extremo anterior é redondeado ou terminado en punta. Presentan un único ollo, ollo nauplio, en posición media, característica típica e conspicua da maioría dos copépodos. Tamén son conspicuas as primeiras antenas unirrámeas (non divididas), que son xeralmente longas e están estendidas en ángulo recto respecto ó eixo lonxitudinal do corpo.
A cabeza está fusionada co primeiro dos seis segmentos torácicos e ás veces tamén co segundo. O primeiro par de apéndices torácicos modificouse en forma de maxilípedos ó servizo da alimentación. Os cinco apéndices torácicos restantes, a excepción do último ou dos últimos pares, son máis ou menos parecidos e birrámeos (divididos en dous), de forma bastante simétrica. Os membros dereito e esquerdo de cada par de apéndices torácicos están unidos entre si por unha placa exoesquelética entre as articulacións coxais. Esta unión permite que as dúas patas batan simultaneamente.
O abdome está composto de cinco segmentos, que son polo xeral máis estreitos que os torácicos. Non hai apéndices abdominais a excepción dun único par de ramas caudais no telson. Nalgunhas especies planctónicas mariñas estas ramas están desenvolvidas de forma espectacular. En Calocalanus pavo, por exemplo, cada rama está xirada lateralmente e porta catro longas sedas en forma de pluma.
Aínda que a maioría dos copépodos son pálidos e transparentes, algunhas especies poden ser dunha cor vermella brillante, laranxas, moradas, azuis ou negras. Coñécense moitas especies luminiscentes.
Seguindo a clasificación taxonómica do CSIC[1], a clase Copepoda divídese en oito ordes:
Outros autores inclúen os branquiúros entre os copépodos, pero o CSIC considera os Branchiura como clase á parte.
Das cinco ordes viventes de copépodos, os calonoideos son maiormente planctónicos; os harpacticoideos, que inclúen arredor do 50% das especies de copépodos, son bentónicos; e os ciclopoideos abarcan especies tanto planctónicas coma epibentónicas.
As variacións con respecto á forma do corpo están relacionadas co hábitat das distintas especies. As formas planctónicas teñen corpos máis ou menos cilíndricos cun abdome estreito. Aquelas que viven en niveis superiores da columna de agua son algo máis miúdas e fusiformes que as que nadan a algúns metros por enriba do fondo. As especies epibentónicas que reptan e nadan xusto por riba do fondo, teñen corpos algo máis anchos. As especies bentónicas que viven sobre as algas e herbas mariñas poden ser anchas e planas; as intersticiais son estreitas e con forma de verme.
O fitoplancto constitúe a parte principal da dieta da maioría das especies, pero algunhas dependen tamén en gran maneira das partículas de detritos. Non tódolos copépodos planctónicos son herbívoros de alimentación suspensívora; algúns son omnívoros, e outros estritamente depredadores. Algunhas especies de Anomalocera y de Pareuchaeta, chegan mesmo a capturar peixes pequenos.
Sábese que as especies de Tisbe, harpacticoideos planctónicos epibentónicos, agrúpanse sobre un peixe pequeno e comen as súas aletas, inmovilizándoo, para logo devora-lo corpo a medida que cae cara o fondo. Algunhas especies dulciacuícolas de Cyclopidae son herbívoras, outras son carnívoras. Os membros do xénero Cyclops son predadores, como son os doutros xéneros dos ciclopoideos.
Nos copépodos de vida libre non hai branquias, e excepto nos calanoideos e nalgunhas especies parasitas non hai nin corazón nin vasos sanguíneos; os órganos excretores son glándulas maxilares.
Os copépodos macho son polo xeral máis pequenos que as femias, que á vez son moito máis abundantes. Os copépodos están entre os poucos crustáceos pequenos que forman espermatóforos, e a parte inferior do espermiducto está modificada con esta finalidade. O xenoporo masculino localízase no primeiro segmento abdominal da maioría dos copépodos, do mesmo xeito que os gonóporos femininos se abren ós receptáculos seminais. Durante a transferencia dos espermatóforos, o macho suxeita á femia cunha ou as dúas primeiras antenas e, na maioría dos calanoideos, tamén co último par de apéndices torácicos modificados. Os espermatóforos son transferidos cos apéndices torácicos do macho e adhírense ás aberturas do receptáculo seminal por medio dun cemento especial.
A maioría dos calanoideos libera os seus ovos individualmente dentro da agua. Porén, os ovos doutros copépodos acostuman a estar pechados nun ovisaco, producido por secrecións do oviduto cando os ovos saen deste, e permanece unido ó segmento xenital feminino, onde funciona a modo de cámara incubadora. Fórmanse un ou dous sacos dependendo do número de ovidutos. Cada saco contén desde uns poucos a 50 ovos ou máis.
Os copépodos son un numerosísimo grupo de pequenos crustáceos, normalmente de menos de 1 mm de lonxitude, pero que representan ó redor do 80% do zooplancto mariño.
A maioría dos copépodos son mariños, pero hai moitas especies dulciacuícolas e unhas poucas que viven entre a vexetación húmida e nas películas de auga na terra. Tamén hai moitas especies parasitas de diferentes animais mariños e dulciacuícolas, especialmente de peixes.
Veslonošci (Kopepodni račići; lat. Copepoda), podrazred sičušnih račića jedva vidljivih golom oku. Narastu 1 - 2 milimetra, a najveći među njima pripadaju rodu Pennella. Mnoge vrste žive sa planktonom i morske su životinje, ali ih se može naći i na vlažnim kopnenim staništima i podzemnim pećinama.
Kopepodni račići važni su u hranidbenom lancu morskih životinja, kao hrana riba, sisavaca i ptica, tako je vrsta Calanus hyperboreus važna je u prehrani jedne male vrste njorke Alle alle. No neke vrste kopepoda žive i kao paraziti na ribama i morskim sisavcima.
Kopepodi su nekada bili klasificirani danas nepriznatom razredu Maxillopoda čije su vrste podijeljene danas na dva nadrazreda, Oligostraca i Multicrustacea, kojemu pripadaju i rakovi Copepoda.
Kopepodni rakovi podijeljeni su na podrazrede Neocopepoda, Progymnoplea i Copepoda incertae sedis sa 21 rodom[1].
Veslonošci (Kopepodni račići; lat. Copepoda), podrazred sičušnih račića jedva vidljivih golom oku. Narastu 1 - 2 milimetra, a najveći među njima pripadaju rodu Pennella. Mnoge vrste žive sa planktonom i morske su životinje, ali ih se može naći i na vlažnim kopnenim staništima i podzemnim pećinama.
Kopepodni račići važni su u hranidbenom lancu morskih životinja, kao hrana riba, sisavaca i ptica, tako je vrsta Calanus hyperboreus važna je u prehrani jedne male vrste njorke Alle alle. No neke vrste kopepoda žive i kao paraziti na ribama i morskim sisavcima.
Kopepodi su nekada bili klasificirani danas nepriznatom razredu Maxillopoda čije su vrste podijeljene danas na dva nadrazreda, Oligostraca i Multicrustacea, kojemu pripadaju i rakovi Copepoda.
Kopepodni rakovi podijeljeni su na podrazrede Neocopepoda, Progymnoplea i Copepoda incertae sedis sa 21 rodom.
Copepoda adalah grup crustacea kecil yang dapat ditemui di laut dan hampir di semua habitat air tawar dan mereka membentuk sumber terbesar protein di samudra.[1] Banyak spesies adalah plankton, tetapi banyak juga spesies benthos dan beberapa spesies kontinental dapat hidup di habitat limno-terestrial dan lainnya di tempat terestrial basah, seperti rawa-rawa.
Copepoda adalah grup crustacea kecil yang dapat ditemui di laut dan hampir di semua habitat air tawar dan mereka membentuk sumber terbesar protein di samudra. Banyak spesies adalah plankton, tetapi banyak juga spesies benthos dan beberapa spesies kontinental dapat hidup di habitat limno-terestrial dan lainnya di tempat terestrial basah, seperti rawa-rawa.
I Copepodi (Copepoda, dal greco kope = remo e podos = piede) sono un raggruppamento di molti piccoli crostacei (raramente più lunghi di 1-2 mm), presenti sia nelle acque marine che in quasi tutti gli habitat dulciacquicoli. Essi rappresentano la più grande fonte di proteine presente negli oceani[1]. Sono i crostacei più numerosi come numero di specie dopo i decapodi e di gran lunga i più numerosi come individui.
I copepodi di solito hanno corpo cilindrico od ovale, con un unico occhio centrale (non sempre presente). Le antenne sono grandi, quelle del primo paio sono molto lunghe e vengono usate per il nuoto. Il torace possiede arti biramificati, l'addome spesso è sottile e non ha appendici. Il corpo termina con alcune appendici o setole terminali. In molte specie le femmine trasportano le uova fecondate in sacche ovigere (una o due a seconda degli ordini) molto vistose
Le specie parassite spesso sono talmente modificate in seguito al loro stile di vita da non avere più alcuna somiglianza con i copepodi a vita libera e, spesso, nemmeno a dei crostacei o a degli animali.
I Copepodi sono esclusivamente acquatici. Questa è però l'unica limitazione: in mare popolano tutti gli habitat, dalle pozze di scogliera alle fosse abissali, e dalle acque polari a quelle equatoriali; popolano poi ambienti dulciacquicoli di diversi tipi, compresi laghi, paludi, sorgenti, acque termali, fiumi e laghi delle grotte, tappeti di muschio, strati di foglie autunnali del sottobosco, acque di fusione dei ghiacciai, pozze piovane formate nelle cavità di piante come le Bromeliacee (fitotelmi).
La maggioranza delle specie sono planctoniche, ma molte sono bentoniche. Costituiscono un anello importantissimo della catena trofica di mari ed oceani.
In maggioranza i Copepodi si nutrono di fitoplancton, soprattutto diatomee. Esistono però numerosissime eccezioni, ad esempio alcuni ordini e famiglie di copepodi (per un totale di circa il 25% delle specie esistenti) sono parassiti di altri animali acquatici.
I popolamenti di questi organismi vengono spesso usati come bioindicatori.
La loro importanza consiste principalmente nel ruolo essenziale che essi hanno come prede in numerosi ecosistemi sia marini che dulcacquicoli consentendo lo sviluppo ed il mantenimento di stock di pesci di importanza commerciale. A livello sperimentale, è stato avviato dal comune di Bologna un progetto per l'utilizzo dei copepodi nell'eliminazione delle larve di zanzara da alcuni bacini idrici. Secondo Marco Farina, curatore del progetto stesso, l'impiego di questi crostacei ha consentito un abbattimento di dette larve superiore al 90%.[2]
I Copepodi costituiscono una sottoclasse (per alcuni studiosi una classe) del subphylum dei Crostacei.
Comprendono oltre 2000 generi e almeno 14000 specie, ripartiti in 10 ordini:
I Copepodi (Copepoda, dal greco kope = remo e podos = piede) sono un raggruppamento di molti piccoli crostacei (raramente più lunghi di 1-2 mm), presenti sia nelle acque marine che in quasi tutti gli habitat dulciacquicoli. Essi rappresentano la più grande fonte di proteine presente negli oceani. Sono i crostacei più numerosi come numero di specie dopo i decapodi e di gran lunga i più numerosi come individui.
Copepoda sunt crustacea classis maxillopodorum.
Copepoda sunt crustacea classis maxillopodorum.
Irklakojai vėžiagyviai (kopepodai) – savarankiški vėžiagyviai, turintys dviskiltį kiautą, kuris užsidaro su atitraukiamojo raumens pagalba. Neturi daugiau kaip dviejų ar trijų liemens išaugų. Gyvena tvenkiniuose ir jūrose, daugiausia dugne. Gėluosiuose vandenyse paplitę kiautavėžiai, mintantys įvairiu maistu.
Planktoniniai kopepodai yra svarbūs ekologiškai bei anglies ciklui, taip pat sudaro didelę dalį žuvų, banginių, jūros paukščių ir kitų vėžiagyvių maisto.
Airkājvēži, peldkājvēži (Copepoda) ir žokļkājvēžu klases apakšklase. Reizēm tā tiek iedalīta kā kārta.
Brīvi dzīvojošie airkāji ir sīki, slaidi, parasti 0,5 - 2 mm gari, retumis līdz 1 cm gari. To ķermenis labi segmentēts; galvas nodalījums saaudzis ar vienu vai diviem krūšu segmentiem. Tiem ir tikai nauplija acs, saliktās acis visiem izzudušas. Līdz ar pieciem divzarainiem, saplacinātiem krūšu kāju pāriem speciāla lokomotoro orgānu funkcija ir arī galvas ekstremitātēm. Vēders parasti sastāv no 4 posmiem. Pirmajā posmā atrodas dzimumatvere. Vēdera nodalījumam nav ekstremitāšu, tā galā ir anālā daiva ar divzarainu dakšiņu. Parazītisko airkāju ķermeņa forma ir ļoti daudzveidīga - gan maisveida, gan ārkārtīgi izstiepta; segmentācija gan vairāk, gan mazāk izzudusi; dakša reducējusies.
Airkāju tēviņi ir mazāki par mātītēm. Pārošanās laikā tēviņš ar taustekļiem satur mātīti un piestiprina tās dzimumatveres tuvumā desveidīgus spermatoforus, no kuriem sperma iekļūst mātītes sēklas uztvērējos un olu dēšanas laika tās apaugļo. Mātīte dažkārt dēj olas katru atsevišķi, bet parasti tās ar olvada sekrētu salipina kopā un piestiprina pie olvada atveres, līdz izšķiļas mazuļi. Airkāji vienmēr attīstās ar metamorfozi. Tā sākas ar nauplija fāzi, kurai seko metanauplija un kopepodīta fāze.
Parazītiskajiem airkājiem no olas kāpurs izšķiļas metanauplija fāzē. Kopepodīta fāzē parazīti piestiprinās pie plekstu dzimtas (Pleuronectidae) zivju žaunām ar pieres dziedzera sekrētu. Kopepodīta fāzē vairākkārt met ādu, līdz ar to regresīvi mainās parazīta forma, turklāt kājas pārvēršas par neposmainiem un nekustīgiem izaugumiem; šādas kopepodīta fāzes stadijas sauc par kūniņu. Pēdējā kūniņas stadija pēc ādas nomešanas pāriet dzimumgatavības fāzē. Šajā fāzē vēži kādu laiku dzīvo brīvi un pārojas. Pēc pārošanās tēviņš nobeidzas, bet mātīte piestiprinās pie jauna saimnieka; šis otrais saimnieks ir kāda mencu dzimtas zivs. Mātītei, kas piestiprinājusies pie zivs žaunām, ķermeņa priekšgalā parādās trīs zaraini izaugumi, kas ieaug saimnieka audos; sakarā ar dzimumdziedzeru milzīgo attīstību vēža tārpveida vēders sasniedz milzīgu apmēru.
Starp airkājiem ir visas pārejas no brīvi dzīvojošām formām līdz tipiskiem parazītiem, par kuru piederību pie airkāju kārtas liecina tikai to attīstība. Daži brīvi dzīvojošie airkāji sastopami atklātos ūdeņos, kur tie ir svarīga un pastāvīga planktona sastāvdaļa. Daļa airkāju sugu ziemā un pavasarī noteikti dominē starp planktona elementiem. Citi turpretim pieder pie piedibens faunas. Jūrās sastop vairāk airkāju sugu nekā saldūdeņos. Ar airkājiem barojas dažādi ūdensdzīvnieki.
Airkājiem ir sastopams paaudžu cikliskums, turklāt atkarībā no klimatiskajiem apstākļiem un ūdensbaseina stāvokļa viena un tā pati suga var būt gan monocikliska, gan dicikliska, gan policikliska. Kā apkārtējie apstākļi ietekmē paaudžu maiņu, par piemēru var noderēt Diaptomus gracilis, kura paaudžu cikls pārtraucas rudenī, ja ūdensbaseins stipri aizaug un pelaģiskais dzīves veids kļūst neiespējams. Neaizaugušā ūdensbaseina Diaptomus gracilis paaudžu monocikliska attīstība notiek cauru gadu.
Pasaulē ir zināmas ap 14000 sugām, kas iedalītas 210 dzimtās un 10 kārtās. Latvijā ir zināmi vairāk, kā 50 sugu pārstāvji no 3 kartām: Ciklopoīdi (Cyclopoida), Kalanoīdi (Calanoida) un Harpaktikoīdi (Harpacticoida).
Apakšklase Airkājvēži (Copepoda)
Airkājvēži, peldkājvēži (Copepoda) ir žokļkājvēžu klases apakšklase. Reizēm tā tiek iedalīta kā kārta.
Eenoogkreeftjes of roeipootkreeftjes (Copepoda) vormen een groep kleine kreeftachtigen. De soorten komen wereldwijd voor en kunnen zowel in zout als zoet water algemeen voorkomen. Er zijn meer dan 13.000 soorten beschreven, waarvan 2800 in zoet water voorkomende soorten. Een bekende groep zijn de in zoet water levende roeipootkreeftjes uit het geslacht Cyclops, die algemeen voorkomen in sloten.[1]
Eenoogkreeftjes hebben net als alle kreeftachtigen een kop, borststuk en achterlijf, waarbij de kop en het borststuk zijn vergroeid. Dit kopborststuk is bij de eenoogkreeftjes bol van vorm. Het staartachtige achterlijf is meer langwerpig van vorm.[1] Eenoogkreeftjes lijken sprekend op elkaar. Ze verschillen van elkaar in de details, zoals het aantal rijen haarachtige structuren op de verschillende delen van het lichaam of de vorm van de pootjes.
Veel soorten zijn vrijzwemmend en leven van diertjes of planten.[1] Andere leven parasitair op gewervelde dieren zoals vissen. Parasitaire eenoogkreeftjes hebben vaak sterk aangepaste lichaamsvormen.
Veel eenoogkreeftjes leven planktonisch (in het water drijvend), maar de meeste leven benthisch (op de bodem). De meeste van de vrijlevende eenoogkreeftjes behoren tot de ordes Calanoida, Cyclopoida en Harpacticoida, alhoewel ook in deze groepen parasitaire vormen voorkomen.
Eenoogkreeftjes of roeipootkreeftjes (Copepoda) vormen een groep kleine kreeftachtigen. De soorten komen wereldwijd voor en kunnen zowel in zout als zoet water algemeen voorkomen. Er zijn meer dan 13.000 soorten beschreven, waarvan 2800 in zoet water voorkomende soorten. Een bekende groep zijn de in zoet water levende roeipootkreeftjes uit het geslacht Cyclops, die algemeen voorkomen in sloten.
Hoppekreps eller copepodar er ei gruppe små krepsdyr som finst i havet og i dei fleste ferskvassmiljø. Dei fleste artane er bentiske, men mange lever òg som plankton og utgjer ein vesentleg del av dyra i planktonet. Somme artar kan òg påtreffast i fuktige miljø på land eller som parasittar på fisk.
Ut frå biomasse og tal på individ kan hoppekreps seiast å vera den mest dominerande dyregruppa på jorda. Det er klassifisert omkring 14 000 artar til no, men det verkelege talet på artar er sannsynlegvis langt høgare.
Dei fleste hoppekreps er mellom 0,5 og 5 mm lange, men kan i kalde farvatn nå lengder på opptil ein cm. Parasittiske artar kan bli endå større: Lernaeocera branchialis, som lever i gjellene til torsk, kan bli heile 3 cm lang.
Som andre krepsdyr har copepodane eit pansra eksoskjelett, men sidan dei er så små er dette som oftast gjennomsiktig i likskap med resten av kroppen. Hoppekreps har tåreforma kropp med lange antenner på over 17 ledd, og mange artar er einaugde.
Ulike planktoniske hoppekreps-artar er særs viktige for så godt som alle marine økosystem som finst. Dei er særs dominerande som planktondyr, og er såleis viktig føde for små fisk, kval, sjøfugl, kril med meir. Dei er òg viktige organismar i karbonkrinsløpet; djuphavet får viktige karbontilskot frå søkkande hoppekrepsskal og -ekskrement.
Parasittiske artar hoppekreps er særs økonomisk skadeleg for fiskeri- og oppdrettsnæringa. M.a. høyrer den berykta lakselusa til gruppa.
Hoppekreps eller copepodar er ei gruppe små krepsdyr som finst i havet og i dei fleste ferskvassmiljø. Dei fleste artane er bentiske, men mange lever òg som plankton og utgjer ein vesentleg del av dyra i planktonet. Somme artar kan òg påtreffast i fuktige miljø på land eller som parasittar på fisk.
Ut frå biomasse og tal på individ kan hoppekreps seiast å vera den mest dominerande dyregruppa på jorda. Det er klassifisert omkring 14 000 artar til no, men det verkelege talet på artar er sannsynlegvis langt høgare.
Hoppekreps eller copepoder (Copepoda) er en klasse av små krepsdyr som lever i havet og i ferskvann. Hoppekrepsene danner en tallrik klasse av krepsdyr, med ca. 13 000 arter i ti ordener. En stor andel av dyreplanktonet i norske hav og andre havområder er hoppekreps; de er viktige beitedyr for fisk, hval og andre krepsdyr som krill. Ved siden av krill er hoppekrepsene den dyregruppa som har den største samla biomassen på jorda. Noen arter er parasitter, og fester seg til fisk, muslinger planter eller koraller. I nordlige farvann er artene av Calanus, spesielt raudåte, ishavsåte og feitåte, svært viktige for økosystemet i havet.[1] Disse kan bli inntil 10 mm lange, mens andre arter som regel bare er noen få millimeter store, som den 1 mm lange Tisbe furcata.[2] Hoppekrepsene er kjøttetere eller altetere som kan beite på planteplankton, mindre dyreplankton eller dødt organisk materiale alt etter mattilgangen.
Hoppekrepser oppstod i havet, men har kolonisert ferskvann ved minst 22 ulike tilfeller. De finnes i alle ferskvannshabitater fra store, gamle innsjøer til underjordiske vannstrømmer, fra dammer av smeltevann til varme kilder, og fra saltsjøer til phytotelma. Noen arter lever på land i myrer og sumper eller i strølaget i fuktige skoger. I ferskvann finnes det over 2800 arter, og artsrikdommen er størst i Palearktis, som har mer enn 1200 arter. Likevel er utbredelsen i saltvann langt større, med minst 11.000 arter.
Marinbiologer ved Gøteborgs Universitet talte på 1990-tallet totalt 1817 arter av hoppekreps i nordlige atlantiske farvann, det vil si på europeisk side av Atlanterhavet inkludert Middelhavet, Østersjøen og Barentshavet.[3] I Bajkalsjøen lever det over 120 arter, og Tanganyikasjøen har 69 arter.[4]
Fiskelus (Caligus curtus), skottelus (Caligus elongatus), lakselus (Lepeophtheirus salmonis) og skrubbelus (Lepeophtheirus perctralis) er eksempler på fiskeparasitter i nordlige farvann. Av andre arter i Norden kan nevnes brislinglus (Lernaeeniscus aprattae), skrubbe-knutecopepode (Acanthocondria cornuta), torske-gjelleorm (Lernaeocera branchialis), og torske-køllecopepode (Clavella adunca).[5]
Hoppekreps er særkjønna. Eggene bæres enten i en eller to eggposer under magen på hunnen, eller de gytes direkte i vannet. Eggene klekkes til naupliuslarver. Som andre krepsdyr vokser hoppekrepsene ved å skifte skall. Larvene utvikler seg gjennom seks stadier, nauplius I-VI. Etter naupliusstadium VI utvikler de seg videre til full voksen størrelse gjennom fem nye stadier. Voksen størrelse for de ulike artene ligger mellom 0,5 og 10 mm.
Lengden på utviklingsstadiene og generasjonslengden er ulik for ulike slekter og arter. For samme art kan generasjonslengden variere med vanntemperaturen; varmere vann gir raskere utvikling og kortere livslengde. Små former kan ha flere generasjoner i løpet av året, mens den vanligste arten i norske farvann, raudåte (Calanus finmarchicus), har én generasjon pr. år. Dens større slektning, Calanus hyperboreus, har en generasjonslengde på to-tre år i arktiske farvann.
Hoppekreps deles i ti ordener (se taksoboks). Av disse er calanoider, cyclopoider og harpacticoider de viktigste i norsk marin fauna. Taksonomien til Maximillopoda er komplisert og under har vært revisjon ettersom ny innsikt vinnes. Det er generelt omstridt å fin-inndele organismer taksonomisk. En vanlig oppdeling anerkjenner Maxllopoda som en av seks ulike klasser av krepsdyr, der forøvrig storkreps (Malacostraca) er den mest artsrike gruppen - etterfulgt av Maxillopoda. En moderne oppdatering av systematikken gis av Martin og Davis[6], som følgende oversikt følger ned til nivået orden, mens lavere nivåer i enkelte tilfeller følger Catalogue of Life: [7]
Hoppekreps eller copepoder (Copepoda) er en klasse av små krepsdyr som lever i havet og i ferskvann. Hoppekrepsene danner en tallrik klasse av krepsdyr, med ca. 13 000 arter i ti ordener. En stor andel av dyreplanktonet i norske hav og andre havområder er hoppekreps; de er viktige beitedyr for fisk, hval og andre krepsdyr som krill. Ved siden av krill er hoppekrepsene den dyregruppa som har den største samla biomassen på jorda. Noen arter er parasitter, og fester seg til fisk, muslinger planter eller koraller. I nordlige farvann er artene av Calanus, spesielt raudåte, ishavsåte og feitåte, svært viktige for økosystemet i havet. Disse kan bli inntil 10 mm lange, mens andre arter som regel bare er noen få millimeter store, som den 1 mm lange Tisbe furcata. Hoppekrepsene er kjøttetere eller altetere som kan beite på planteplankton, mindre dyreplankton eller dødt organisk materiale alt etter mattilgangen.
Hoppekrepser oppstod i havet, men har kolonisert ferskvann ved minst 22 ulike tilfeller. De finnes i alle ferskvannshabitater fra store, gamle innsjøer til underjordiske vannstrømmer, fra dammer av smeltevann til varme kilder, og fra saltsjøer til phytotelma. Noen arter lever på land i myrer og sumper eller i strølaget i fuktige skoger. I ferskvann finnes det over 2800 arter, og artsrikdommen er størst i Palearktis, som har mer enn 1200 arter. Likevel er utbredelsen i saltvann langt større, med minst 11.000 arter.
Marinbiologer ved Gøteborgs Universitet talte på 1990-tallet totalt 1817 arter av hoppekreps i nordlige atlantiske farvann, det vil si på europeisk side av Atlanterhavet inkludert Middelhavet, Østersjøen og Barentshavet. I Bajkalsjøen lever det over 120 arter, og Tanganyikasjøen har 69 arter.
Fiskelus (Caligus curtus), skottelus (Caligus elongatus), lakselus (Lepeophtheirus salmonis) og skrubbelus (Lepeophtheirus perctralis) er eksempler på fiskeparasitter i nordlige farvann. Av andre arter i Norden kan nevnes brislinglus (Lernaeeniscus aprattae), skrubbe-knutecopepode (Acanthocondria cornuta), torske-gjelleorm (Lernaeocera branchialis), og torske-køllecopepode (Clavella adunca).
Widłonogi (Copepoda) – drobne, z reguły planktonowe, morskie i słodkowodne skorupiaki. Zasiedlają wszystkie oceany i kontynenty łącznie z Antarktydą. Występują wszędzie tam, gdzie jest woda, nawet w wilgotnym mchu, pod korą dolnych części pni drzew tropikalnych, w wilgotnym piasku plaż morskich. Część widłonogów prowadzi pasożytniczy tryb życia, pasożytując na jamochłonach, pierścienicach, skorupiakach, mięczakach, szkarłupniach, rybach. Ciało jest zbudowane ze stałej liczby 16 segmentów (5 głowowych, 6 tułowiowych i 5 odwłokowych). Rozmiar ich ciała zawiera się w zakresie od 0,5 mm do 20 cm, jednak typowi przedstawiciele tej podgromady mają zwykle rozmiar ciała zawierający się w zakresie 0,5 do 2 mm. Pływają za pomocą silnie rozwiniętej pierwszej pary czułków. Ciało ich zawiera żółte kropelki zapasowego tłuszczu (glikolipidy) oraz wiele witamin. Przeważnie są przezroczyste. W morzach stanowią około 9/10 planktonu. Są też głównym składnikiem pokarmu dla śledzi i narybku wątłuszowego (Gadidae). Również gatunki słodkowodne odgrywają pewną rolę jako pokarm dla ryb. Wolno żyjące oraz pasożytnicze widłonogi są rozdzielnopłciowe. Jaja u wielu widłonogów składane są do worków lęgowych znajdujących się po bokach odwłoka. Widłonogi stanowią dużą grupę składającą się obecnie z 9 rzędów, około 220 rodzin i ponad 13 000 gatunków, co czyni je najliczniejszą gromadą wśród skorupiaków niższych.
Podgromada widłonogi dzieli się na następujące rzędy:
Ze względu na różnorodność gatunkową, gromadę tę cechuje duże zróżnicowanie morfologiczne. Łączy je jednak stała liczba segmentów ciała, brak karapaksu, występowanie na końcu odwłoka charakterystycznych widełek (skąd nazwa gromady), noszenie przez samice jaj przyklejonych do ciała[1].
Ze złożonych przez samice jaj wykluwają się larwy. Przechodzą one 10-11 linień. Pierwsze stadia larwalne to naupliusy lub metanaupliusy (5-6 linień). Kolejne stadia larwalne to kopepodity (następne 5 linień). U form dojrzałych występuje wyraźny dymorfizm płciowy. Samce są mniejsze. Kilka gatunków pasożytów jest obojnaczych. Partenogeneza występuje tylko w rzędzie Harpacticodia[1].
Widłonogi (Copepoda) – drobne, z reguły planktonowe, morskie i słodkowodne skorupiaki. Zasiedlają wszystkie oceany i kontynenty łącznie z Antarktydą. Występują wszędzie tam, gdzie jest woda, nawet w wilgotnym mchu, pod korą dolnych części pni drzew tropikalnych, w wilgotnym piasku plaż morskich. Część widłonogów prowadzi pasożytniczy tryb życia, pasożytując na jamochłonach, pierścienicach, skorupiakach, mięczakach, szkarłupniach, rybach. Ciało jest zbudowane ze stałej liczby 16 segmentów (5 głowowych, 6 tułowiowych i 5 odwłokowych). Rozmiar ich ciała zawiera się w zakresie od 0,5 mm do 20 cm, jednak typowi przedstawiciele tej podgromady mają zwykle rozmiar ciała zawierający się w zakresie 0,5 do 2 mm. Pływają za pomocą silnie rozwiniętej pierwszej pary czułków. Ciało ich zawiera żółte kropelki zapasowego tłuszczu (glikolipidy) oraz wiele witamin. Przeważnie są przezroczyste. W morzach stanowią około 9/10 planktonu. Są też głównym składnikiem pokarmu dla śledzi i narybku wątłuszowego (Gadidae). Również gatunki słodkowodne odgrywają pewną rolę jako pokarm dla ryb. Wolno żyjące oraz pasożytnicze widłonogi są rozdzielnopłciowe. Jaja u wielu widłonogów składane są do worków lęgowych znajdujących się po bokach odwłoka. Widłonogi stanowią dużą grupę składającą się obecnie z 9 rzędów, około 220 rodzin i ponad 13 000 gatunków, co czyni je najliczniejszą gromadą wśród skorupiaków niższych.
Podgromada widłonogi dzieli się na następujące rzędy:
Os copépodes (subclasse Copepoda) são um grupo de crustáceos muito importante na composição da fauna de invertebrados aquáticos. Há cerca de 12000 espécies conhecidas; dessas, 7500 são de vida livre, sendo 1200 próprias de águas continentais.[1]
Certos copépodes de águas continentais podem formar agregações com densidades superiores a 11000 indivíduos por litro. Os copépodes podem parasitar peixes e invertebrados aquáticos.
A subclasse Copepoda é a maior e mais diversificada dos crustáceos. Além disso, é considerado o grupo de organismos pluricelulares mais abundante no planeta, superando em número de indivíduos até os insetos. Esses microcrustáceos habitam os diversos ambientes aquáticos, incluindo terras (h)úmidas. Os crustáceos de vida livre podem ser planctônicos, bentônicos, viver em águas subterrâneas, entre outros ambientes. Os copépodes de águas continentais não são tão diversos como os de água salgada, sendo que o plâncton é composto principalmente por Calanoida e Cyclopoida.
A maioria dos copépodes possuem de 1 a 5 mm de comprimento. Seu corpo é composto de cabeça, tórax e abdômen, sendo sua extremidade anterior arredondada ou pontiaguda. A cabeça está fundida com o primeiro e às vezes o segundo segmento torácico. Não possuem olhos compostos, porém há um olho naupliano mediano típico na maioria. As primeiras antenas são unirremes e longas. O tórax possui seis segmentos. O primeiro par de apêndices é modificado, formando maxilípedes para alimentação. Os outros cinco pares de apêndices são semelhantes e simetricamente birremes. Entretanto, em algumas espécies, os apêndices do último par, conhecido como P5, podem vir modificados para a cópula, assimétricos ou até mesmo unirremes. O abdômen possui cinco segmentos que são mais estreitos que o do tórax e não possui apêndices, exceto um par de ramos caudais no télson.
Esta espécie microscópica é a mais rápida do planeta, conseguindo percorrer por segundo uma distância equivalente a 500 vezes o seu comprimento, atingindo os 11 km/h. Comparando com a chita, que é o animal terrestre mais veloz, esta teria de ser capaz de atingir 8000 km/h para ser tão rápida como o copépode. O professor Thomas Kiørboe, do Instituto Nacional de Recursos Aquáticos na Universidade da Dinamarca, fez uma filmagem para determinar e mostrar a velocidade do salto de escape do animal ao se sentir ameaçado, sendo esta meio metro por segundo.
Além de mais rápidos, são considerados os animais mais fortes do mundo, sendo de 10 a 30 vezes mais fortes do que qualquer outro animal, considerando proporções de tamanho.
Os copépodes possuem dois sistemas motrizes, formados por 4 a 5 pares de patas para nadar e saltar, além de um sistema de extremidades que vibra para ajudar na continuidade do movimento enquanto nada. Sua forma física é otimizada para altas velocidades e seus diversos sistemas de movimento adicionados a extremidade vibratória faz com que o animal, além de forte e veloz, não sinta fadiga.
Em aquários marinhos, os copépodes podem ser observados a olho nu em seu período de maturação, se apresentando como pequenos pontos brancos em movimento na água e no vidro. A alimentação desses animais é à base de pequenas algas, ou fitoplâncton, e portanto se o aquário possuir boa higienização e iluminação, os copépodes permanecerão neles até que se ponha um peixe no aquário, que se alimentará deles. Se o aquário já possui peixes e aparecem copépodes, provavelmente será quando uma nova água for posta no aquário.
Apenas uma espécie de copépode, o Ergasilus sieboldi, poderá trazer problemas ao peixe, fixando-se em suas brânquias ou até mesmo olhos. Ao atacar um peixe, esse copépode causa sua degeneração por hemorragia e necrose e essa infestação pode ser notada pela dificuldade de respiração do peixe e sua instalação pode ser confirmada com a observação através de uma lupa, não sendo o Ergasilus sieboldi possível de ver a olho nu.
Tratamentos como banhos de formalina ou remédios a base de cobre são eficientes, juntamente com um cuidado especial para com o peixe em questão, que pode livrar-se do problema usando suas defesas naturais contra o parasita.
Ordens de copépodes:
Os copépodes (subclasse Copepoda) são um grupo de crustáceos muito importante na composição da fauna de invertebrados aquáticos. Há cerca de 12000 espécies conhecidas; dessas, 7500 são de vida livre, sendo 1200 próprias de águas continentais.
Certos copépodes de águas continentais podem formar agregações com densidades superiores a 11000 indivíduos por litro. Os copépodes podem parasitar peixes e invertebrados aquáticos.
A subclasse Copepoda é a maior e mais diversificada dos crustáceos. Além disso, é considerado o grupo de organismos pluricelulares mais abundante no planeta, superando em número de indivíduos até os insetos. Esses microcrustáceos habitam os diversos ambientes aquáticos, incluindo terras (h)úmidas. Os crustáceos de vida livre podem ser planctônicos, bentônicos, viver em águas subterrâneas, entre outros ambientes. Os copépodes de águas continentais não são tão diversos como os de água salgada, sendo que o plâncton é composto principalmente por Calanoida e Cyclopoida.
A maioria dos copépodes possuem de 1 a 5 mm de comprimento. Seu corpo é composto de cabeça, tórax e abdômen, sendo sua extremidade anterior arredondada ou pontiaguda. A cabeça está fundida com o primeiro e às vezes o segundo segmento torácico. Não possuem olhos compostos, porém há um olho naupliano mediano típico na maioria. As primeiras antenas são unirremes e longas. O tórax possui seis segmentos. O primeiro par de apêndices é modificado, formando maxilípedes para alimentação. Os outros cinco pares de apêndices são semelhantes e simetricamente birremes. Entretanto, em algumas espécies, os apêndices do último par, conhecido como P5, podem vir modificados para a cópula, assimétricos ou até mesmo unirremes. O abdômen possui cinco segmentos que são mais estreitos que o do tórax e não possui apêndices, exceto um par de ramos caudais no télson.
Esta espécie microscópica é a mais rápida do planeta, conseguindo percorrer por segundo uma distância equivalente a 500 vezes o seu comprimento, atingindo os 11 km/h. Comparando com a chita, que é o animal terrestre mais veloz, esta teria de ser capaz de atingir 8000 km/h para ser tão rápida como o copépode. O professor Thomas Kiørboe, do Instituto Nacional de Recursos Aquáticos na Universidade da Dinamarca, fez uma filmagem para determinar e mostrar a velocidade do salto de escape do animal ao se sentir ameaçado, sendo esta meio metro por segundo.
Além de mais rápidos, são considerados os animais mais fortes do mundo, sendo de 10 a 30 vezes mais fortes do que qualquer outro animal, considerando proporções de tamanho.
Os copépodes possuem dois sistemas motrizes, formados por 4 a 5 pares de patas para nadar e saltar, além de um sistema de extremidades que vibra para ajudar na continuidade do movimento enquanto nada. Sua forma física é otimizada para altas velocidades e seus diversos sistemas de movimento adicionados a extremidade vibratória faz com que o animal, além de forte e veloz, não sinta fadiga.
A aparição dos Copépodes num aquárioEm aquários marinhos, os copépodes podem ser observados a olho nu em seu período de maturação, se apresentando como pequenos pontos brancos em movimento na água e no vidro. A alimentação desses animais é à base de pequenas algas, ou fitoplâncton, e portanto se o aquário possuir boa higienização e iluminação, os copépodes permanecerão neles até que se ponha um peixe no aquário, que se alimentará deles. Se o aquário já possui peixes e aparecem copépodes, provavelmente será quando uma nova água for posta no aquário.
Apenas uma espécie de copépode, o Ergasilus sieboldi, poderá trazer problemas ao peixe, fixando-se em suas brânquias ou até mesmo olhos. Ao atacar um peixe, esse copépode causa sua degeneração por hemorragia e necrose e essa infestação pode ser notada pela dificuldade de respiração do peixe e sua instalação pode ser confirmada com a observação através de uma lupa, não sendo o Ergasilus sieboldi possível de ver a olho nu.
Tratamentos como banhos de formalina ou remédios a base de cobre são eficientes, juntamente com um cuidado especial para com o peixe em questão, que pode livrar-se do problema usando suas defesas naturais contra o parasita.
Copepodele (Copepoda) (din greaca kope = vâslă + latina poda = picior) sunt o subclasă de crustacee inferioare mici (între 0,1 și 2 mm, excepțional până la 10 mm) care cuprinde specii marine și dulcicole, multe parazite. Au corpul alungit, neacoperit de carapace, divizat în segmente mici chitinoase (5 cefalice, 6 toracice și 5 abdominale). Primul segment toracic, adesea și al doilea și chiar mai multe, sunt contopite cu capul. La cap au 2 antenule și 2 antene și dorsal un ochi median nauplian. Antenulele sunt mai dezvoltate, ele jucând un rol important în locomoție, în coordonare cu mișcările picioarelor. Acestea sunt biramate, scurte, turtite în plan transversal și cu o bogată garnitură de peri dispuși în același plan, astfel că formează o bună ramă de înotat , de unde și denumirea de Copepoda. Piesele aparatului bucal care servesc pentru hrană sunt formate de o pereche de mandibule, două perechi de maxile și o pereche de maxilipede. Toracele, format din șase segmente, are 5 perechi de picioare articulate (toracopode) care servesc pentru vâslit. Abdomenul se termină cu o furcă și are peri chitinoși. Au sexe separate. Femelele au primele două segmente abdominale sudate și unul sau doi saci ovigeri. Formele libere se hrănesc cu alge sau cu nevertebrate mici, de exemplu cu larve de chironomide sau de alte nevertebrate, cu ouă sau chiar cu alte copepode. Formele parazite se găsesc atât în apele dulci, unde parazitează numai pe pești, cât și în apele marine, unde parazitează pe pești și pe nevertebrate. Copepodele sunt o parte importantă a planctonului. Prezintă importanță mare pentru piscicultură, fiindcă servesc ca hrană principală pentru pești, iar unele specii sunt redutabili paraziți ai peștilor.
Copepodele (Copepoda) (din greaca kope = vâslă + latina poda = picior) sunt o subclasă de crustacee inferioare mici (între 0,1 și 2 mm, excepțional până la 10 mm) care cuprinde specii marine și dulcicole, multe parazite. Au corpul alungit, neacoperit de carapace, divizat în segmente mici chitinoase (5 cefalice, 6 toracice și 5 abdominale). Primul segment toracic, adesea și al doilea și chiar mai multe, sunt contopite cu capul. La cap au 2 antenule și 2 antene și dorsal un ochi median nauplian. Antenulele sunt mai dezvoltate, ele jucând un rol important în locomoție, în coordonare cu mișcările picioarelor. Acestea sunt biramate, scurte, turtite în plan transversal și cu o bogată garnitură de peri dispuși în același plan, astfel că formează o bună ramă de înotat , de unde și denumirea de Copepoda. Piesele aparatului bucal care servesc pentru hrană sunt formate de o pereche de mandibule, două perechi de maxile și o pereche de maxilipede. Toracele, format din șase segmente, are 5 perechi de picioare articulate (toracopode) care servesc pentru vâslit. Abdomenul se termină cu o furcă și are peri chitinoși. Au sexe separate. Femelele au primele două segmente abdominale sudate și unul sau doi saci ovigeri. Formele libere se hrănesc cu alge sau cu nevertebrate mici, de exemplu cu larve de chironomide sau de alte nevertebrate, cu ouă sau chiar cu alte copepode. Formele parazite se găsesc atât în apele dulci, unde parazitează numai pe pești, cât și în apele marine, unde parazitează pe pești și pe nevertebrate. Copepodele sunt o parte importantă a planctonului. Prezintă importanță mare pentru piscicultură, fiindcă servesc ca hrană principală pentru pești, iar unele specii sunt redutabili paraziți ai peștilor.
Ceponožci ali ceponožni raki (znanstveno ime Copepoda) so številčna skupina majhnih rakov, ki poseljuje morja in oceane ter praktično vse sladkovodne habitate, vključno z začasnimi vodnimi telesi in dovolj vlažnimi prostorčki v mahu, rastlinskem opadu in med listi. Mnoge vrste so tudi jamske.
V splošnem so majhni raki, ki zrastejo v dolžino med 1 in 5 mm, največji zajedavski ceponožci pa so lahko dolgi več deset centimetrov. Največkrat so prozorni, nekatere vrste pa so živih barv ali celo luminiscenčne. Oblika telesa je odvisna od okolja, v katerem posamezna vrsta živi, največkrat pa se njihovo telo stožčasto oža proti zadnjemu delu, glava pa je zaokrožena ali koničasta.
Glava je zraščena s prvim in včasih še drugim členom oprsja. Najopaznejši del glave je največkrat par zelo dolgih, nerazvejanih tipalnic, ki jih ceponožci držijo pravokotno na telo. Njegova vloga je povečevanje plovnosti, zato je poraščen s ščetinami in razvit predvsem pri planktonskih vrstah. To je prvi par, drugi par je krajši in dvovejnat, sodeluje pri plavanju. Med tipalnicami je neparno navplijevo oko. Prvi par oprsnih nožic (torakopodov) je spremenjen v obustne okončine in sodeluje pri prehranjevanju, nadaljnji trije ali štirje so simetrično dvovejnati in so udeleženi pri premikanju, zadnji par ali dva pa sta prilagojena razmnoževanju. Vsak par plavalnih nog je prečno povezan med seboj s ploščo skeleta, tako da se lahko premikajo le simultano.
Zadek, ki je grajen iz petih členov, se proti koncu počasi oža in nima okončin, razen na analnem členu, kjer je par izrastkov. Ta sta lahko različno oblikovana, pri nekaterih planktonskih vrstah celo v spektakularnih oblikah in barvah.
Ceponožci so posebnost med majhnimi raki v tem, da samci tvorijo spermatofor za oploditev samice. Po navadi so manjši in manj številčnejši od samic. Po parjenju samica tvori jajčno vrečko, v katero izleže do 50 ali več jajčec.
Ličinkam pravimo navpliji. Ko se petkrat ali šestkrat levijo, postanejo podobni odraslim živalim, a imajo nečlenjen zadek in manj parov nog. Odrastejo po dodatnih petih levitvah. Celoten stadij ličinke lahko traja od enega tedna do skoraj enega leta, celotna življenjska doba pa znaša od pol leta do malo več kot enega leta pri prosto živečih vrstah.
Hladna obdobja leta preživijo morske vrste kot jajčeca, kopenske vrste pa mirujejo obdana z neprepustnim ovojem in zakopana v blatu ter so sposobna preživeti obdobja, ko se njihovo okolje povsem izsuši.
Večina vrst je morskih, mnogo pa jih poseljuje tudi sladkovodne habitate ali živijo v drobnih z vodo napolnjenih prostorčkih na kopnem. Tudi po načinu prehranjevanja je skupina zelo raznolika. Med njimi so plenilske in rastlinojede vrste, velik delež pa je tudi zajedavcev. Posebej morske vrste so lahko zelo številčne in pogosto predstavljajo največji delež planktona po številu osebkov na določenem območju. Rastlinojede morske vrste, ki se prehranjujejo s fitoplanktonom, so tako pomemben povezovalni element med primarnimi producenti in organizmi višje v prehranjevalni verigi. Mnogim od teh organizmov so poglavitni vir hrane. Nekateri znanstveniki celo ocenjujejo, da predstavljajo največji delež biomase v svetovnih oceanih in jih imenujejo »žuželke morja«.[1]
Planktonski (bentoški) ceponožci so največkrat filtratorji, ki lovijo organski drobir in planktonske alge (npr. diatomeje). Druge vrste so vsejede ali plenilske, nekatere lovijo celo manjše ribe. Ceponožci, ki živijo pri dnu, največkrat jedo mikroorganizme in organski drobir, pritrjen na podlago.
Vsaj tretjina vrst je zajedavskih. Njihovi gostitelji so lahko ribe, mnogoščetinci, iglokožci, plaščarji ali školjke. Nekateri zajedajo tako, da se s preobraženimi nogami pritrdijo na gostitelja in z ostrimi obustnimi okončinami predrejo njegov integument ali škrge. Ti so še razmeroma podobni prostoživečim ceponožcem. Notranji zajedavci pa so pogosto toliko spremenjeni, da ne spominjajo več na ceponožce in jih je možno uvrstiti v to skupino le po ličinkah, ki niso prilagojene zajedavstvu.
Četudi živijo v praktično vseh sladkovodnih habitatih, večinoma ne predstavljajo tveganja za prečiščeno pitno vodo, le v nekaterih tropskih državah so opazili povezavo med njimi in izbruhi kolere. Domnevajo, da ceponožci delujejo kot prenašalci bakterij Vibrio cholerae, ki se oprimejo njihove površine.[2] Po drugi strani se uspešno uporabljajo za biološko kontrolo ličink komarjev, ki prenašajo nevarne nalezljive bolezni, in jih namenoma naseljujejo v rezervoarje za deževnico v Vietnamu.[3]
Nekatere vrste ceponožcev gojijo akvaristi za hrano ribam.
Opisanih je preko 12.000 vrst ceponožcev, s čemer so med številčnejšimi skupinami rakov. Združujemo jih v 10 redov. Za Slovenijo je znanih dobrih 100 sladkovodnih vrst. Za Tržaški zaliv zadnji pregledne raziskave v začetku 20. stoletja navajajo 160 vrst, a se je medtem okolje bistveno spremenilo, tako da o sodobni favni tega območja ni preglednih podatkov.
Ceponožci ali ceponožni raki (znanstveno ime Copepoda) so številčna skupina majhnih rakov, ki poseljuje morja in oceane ter praktično vse sladkovodne habitate, vključno z začasnimi vodnimi telesi in dovolj vlažnimi prostorčki v mahu, rastlinskem opadu in med listi. Mnoge vrste so tudi jamske.
Hoppkräftor (Copepoda) är små kräftdjur, några mm i längd, som förekommer i både hav och sötvatten. En del arter är plankton (frisimmande), en del är bentiska (bottenlevande). De lever som filtrerare, rovdjur eller parasiter. Med 13 000 arter är de den artrikaste gruppen bland kräftdjuren och utgör en huvuddel av marint zooplankton. Dessa skenbart obetydliga kräftdjur är kanske störst i djurriket både beträffande individantal och biomassa och en ytterst viktig del av de marina näringskedjorna. De har avgörande betydelse för bestånden av fisk och högre djur. De har också mycket stor betydelse för kolets kretslopp och därmed klimatet.[1][2][3]
Några kända hoppkräftor är de mm-stora Cyclops som är mycket vanliga i sötvatten, de större Calanus som dominerar i kalla hav och Harpacticider som är små och bottenlevande i haven.
Kroppslängden är vanligen mellan 0,2 och 2 mm. Hos den minsta arten (Sphaeronellopsis monothrix), som lever innanför skalet hos musselkräftor, är hanen är 0,11 mm. Den största arten (Penella balaenoptera), en parasit på bardvalar, blir upp till 320 mm. Kroppsformen är långsträckt till droppformig, men kan vara annorlunda beroende på levnadssättet. Pelagiska arter har mer ursprunglig kroppsform, bottenlevande arter är mera plattade och grävande arter mera cylindriska. De parasitiska arterna är de mest modifierade; hos dem kan man inte se att den vuxna honan är en hoppkräfta, utan detta framgår bara av tidigare utvecklingsstadier. Liksom andra kräftdjur har de ett exoskelett, men detta är tunt och kroppen är nästan genomskinlig. Hoppkräftornas färg beror på upplagrade oljedroppar som används som reservfett. Hudpigment finns sällan.
Kroppen består av huvud och tio kroppssegment, sex thoraxsegment med benpar och fyra abdominalsegment. Huvudet och första thoraxsegmentet är alltid sammansmälta. Sköld (carapax) saknas. Extremiteterna är som hos alla kräftdjur tvågrenade (Y-formade).
Huvudet har två par antenner. Första paret (antennula) är mindre och olika beroende på levnadssättet. Hos Calanoida är de längre än halva kroppslängden, hos Cyclopoida kortare. Andra antennparet är hos frilevande copepoder vanligen det viktigaste organet för förflyttning; de rör sig som åror för att dra djuret genom vattnet. Huvudet har också mundelar, bl.a. mandibler (käkar) och maxiller. Mandiblernas och munnens form beror på levnadssättet och är viktiga taxonomiska kännetecken. Hos gruppen Siphonostomatida är t.ex. labrum och labium (överläpp och underläpp) sammansmälta och ofta utdragna till ett långt sugrör. Oftast finns ett enda sammansatt öga beläget i huvudets mittlinje och ofta klart rött. Grottlevande arter kan sakna öga.
Thorax (mellankroppen) består av sex segment med benpar. Första thoraxbenparet är omvandlat till ogrenade s.k. maxillipeder, som används vid födointag. De följande fyra benparen är tvågrenade simben. Det sjätte benparet är ofta reducerat.
Abdomen (bakkroppen) är typiskt smalare än thorax och består av fyra eller fem segment utan extremiteter. Könsöppningen finns på första abdominalsegmentet, där hos honan en eller två äggsäckar kan vara fästa. Bakkroppen avslutas med en platta (telson) som kan ha svanslika bihang (furca) med betydelse för flytförmåga och styrning.
På grund av sin litenhet behöver copepoderna oftast inget hjärta och inget blodkärlssystem. Vissa arter i ordningen Calanoida har dock ett hjärta, men inga blodkärl, utan öppet kretslopp. De flesta copepoderna saknar också gälar, och tar upp syre direkt genom kroppsytan. Utsöndringen sköts av körtlar nära maxillerna.
Copepoder har många sinnesorgan. De har borstliknande sinneshår som fungerar som mekanoreceptorer och reagerar på strömningar i vattnet, som böjer håren. Därigenom kan de detektera både bytesdjur och rovdjur i omgivningen, och genom att känna igen strömningsmönstret kan de skilja mellan dessa.
Många copepoder visar extremt snabba flyktreaktioner då rovdjur närmar sig, varvid de flyttar sig några millimeter. Många arter har nervceller omgivna av myelin (som ökar ledningshastigheten), vilket är mycket ovanligt bland ryggradslösa djur.
Honor av vissa arter avger feromoner, som bildar ett spår i vattnet som hanen kan följa.
Alla hoppkräftor är skildkönade och har utpräglad könsdimorfism. Hanarna är alltid mindre än honorna och har ofta första antennparet ombildat till ett griporgan, och de utgör ofta mindre än hälften av populationen. Partenogenes finns bara hos ett fåtal arter. Parningen har undersökts hos gruppen Calanoida. Hanen griper med en av sina antenner i första paret (vanligen den högra) tag i honans bakkroppsbihang. Hanen vänder sig sedan så att djuren får motsatt riktning. Med höger 6:e thoraxben fattar han sedan om honans bakkropp och överför med vänster 6:e thoraxben en spermatofor (spermiepaket) till hennes första abdominalsegment. Då äggen läggs bildas äggsäckar som är fästa vid honans bakkropp. Äggen blir kvar där tills naupliuslarverna kläcks. Hos några arter som lever i dammar och andra småvatten har äggen ett kraftigt skal och kan överleva lång tid ifall vattensamlingen torkar ut.
Äggen kläcks nästan alltid till naupliuslarver, som har ett huvud och en svans, men ingen thorax eller egentlig abdomen. Ett undantag är vissa parasiter som har starkt förkortad livscykel och kläcks som copepoditlarver. Naupliuslarven ömsar hud fem eller sex gånger innan den blir en s.k. copepoditlarv. Denna liknar det vuxna djuret, men har en enkel osegmenterad abdomen och bara tre thorakala benpar. Efter ytterligare fem hudömsningar är djuret vuxet. Hela processen från kläckning till vuxen kan ta från en vecka till ett år, beroende på arten.
Livslängden beror starkt av temperaturen. Musselparasiten Mytilicola intestinalis lever t.ex. 3-6 månader i Medelhavet och har flera generationer per år, medan den vid Englands sydkust bara har två generationer under sommaren, och individerna blir 9-12 månader. Stora arter har livslängd 8-14 månader. Som överlevnadsstrategi för ogynnsamma årstider, fr.a. vintern, bildar flera arter motståndskraftiga ägg. Det finns dock även arter som är aktiva under vintern och överlever sommaren genom att bilda cystor.
Hoppkräftornas näring är mångsidig. Alla näringssätt kan dock härledas från ett liv som rovdjur. Det finns filtrerare som äter detritus och alla slags växtplankton, och arter som aktivt griper små näringspartiklar. Många arter gör bådadera. Många bentiska copepoder äter organisk detritus och bakterier och deras mundelar är ombildade för att skrapa eller bita. Herbivora arter kan filtrera alger ner till 5 μm. Karnivora arter filtrerar djur mindre än 30-50 μm, större fångas aktivt. De äter skallösa protozoer, andra hoppkräftor och t.o.m. små fiskyngel, så stora bytesdjur de kan fånga. Bland kommensaler och parasiter ses ett spektrum från enkla avfallsätare till stationära blodsugare. Ekto- och endoparasiter lever på hudflagor, slem osv resp. blod och vävnadsvätska.
Växtätande copepoder, särskilt i näringsrika kalla hav, lagrar energi från födan som oljedroppar under vårens och sommarens planktontoppar. Dessa kan uppgå till halva kroppsvolymen.
Hoppkräftorna har kosmopolitisk utbredning och finns i alla typer av vatten, från oceanerna till de minsta vattensamlingarna. De saknas bara i ständigt istäckta eller vattenlösa områden. Omkring 80% av de nu levande arterna lever i havet, där det största individantalet och artantalet finns i de översta skiktet samt i litoralen. Hoppkräftor har anträffats ned till 5 000 m djup. Några arter har trängt ner i grundvattnet.
Mycket gamla sjöar är hotspots för biodiversitet. Av de 120 arter hoppkräftor som finns i Bajkalsjön är de flesta endemiska. Samma gäller Tanganyikasjön.
Hoppkräftornas miljöer är mycket varierande. Det finns frisimmande (plankton), bottenlevande (bentiska), grävande och parasitiska arter. Det finns arter som lever i våta terrestra miljöer som våtmarker, källor, tillfälliga vattensamlingar, fuktig mossa eller vattenfyllda delar av växter som bromeliacéer eller kannrankeväxter. Många lever i grottor och hålrum i sötvatten eller saltvatten. De livnär sig som filtrerare eller rovdjur.
Hoppkräftorna är förmodligen den individrikaste gruppen bland flercelliga djur på jorden. I övre skikten i nästan alla havsytor är ca 90% av zooplankton hoppkräftor. Eftersom de också ofta uppträder i täta svärmar (ca 10 000 individer / m3 hos Pseudocalanus-arter) har de avgörande betydelse som näring för många fiskarter, inklusive sådana som används av människan. Även i sötvatten är hoppkräftor den viktigaste näringskällan för många fiskar. Å andra sidan kan hoppkräftor skada fiskodlingar i sötvatten genom att angripa yngel. Näringskedjan är mycket kort, då många hoppkräftor lever av växtplankton och sedan direkt äts av fiskar. Döda hoppkräftor bryts snabbt ner: vid 22 °C tar detta 3 dagar.
Copepoder i plankton har stor betydelse för ekosystemen och kolets kretslopp. De är ofta dominerande zooplankton och den viktigaste födan för många fiskar, valar, sjöfåglar och andra kräftdjur som krill. Vissa forskare anser att de utgör den största djurbiomassan på jorden (antarktisk krill är dock jämförbar). Calanus glacialis, som lever vid kanterna av de arktiska isarna, utgör upp till 80% av biomassan i zooplankton där. Populationerna har en topp då isarna drar sig tillbaka på våren. Den nutida minskningen av arktiska isar kan tvinga dem att konkurrera i den öppna oceanen med C. finmarchicus, som är näringsfattigare och nu sprider sig från Nordsjön och Norska havet upp i Barents hav.[4]
På grund av sin litenhet och sin snabba tillväxt, och eftersom de är spridda i alla oceaner, bidrar copepoderna nästan säkert mer till oceanernas sekundärproduktion och kolsänka än krill och kanske mer än alla andra organismer tillsammans. Oceanernas ytskikt anses som världens största kolsänka och tar upp ca 2 miljarder ton kol varje år, motsvarande en tredjedel av mänsklighetens koldioxidutsläpp. Många copepoder äter nära ytan på natten och vistas dagtid på större djup för att undvika rovdjur (de ändrar sin densitet genom att omvandla oljor till tätare fetter). Genom utbytta exoskelett, faeces, döda individer och andning på större djup transporteras kol nedåt i haven.
Omkring hälften av de 13 000 arterna är parasiter. Dessa har starkt modifierad kropp. De fäster sig på fiskar, hajar, marina däggdjur och många ryggradslösa djur som mollusker, tunikater och koraller i haven. De kan också vara både endo- och ektoparasiter på fisk och ryggradslösa djur i sötvatten.
Hoppkräftor (Copepoda) bildar en underklass bland kräftdjuren. Omkring 13 000 arter är kända, varav ca 2800 i sötvatten.
För gruppen hoppkräftor gäller följande autapomorfier:
Huys och Boxshall[5] urskiljer 9 ordningar bland copepoderna, vilkas namn och släktskap framgår av följande kladogram:
Hoppkräftor (Copepoda) N.N.Hoppkräftor används ofta som fiskfoder till akvariefiskar, särskilt i saltvattensakvarier. De gör också nytta genom att förtära avfall och oönskade alger i akvarier.
Hoppkräftor förekommer ibland i dricksvattensystem i städer, särskilt om vattnet inte är filtrerat. Detta har inträffat i New York, Boston och San Francisco. Oftast är detta inget problem om vattnet renas. I tropiska länder som Peru och Bangladesh har man påvisat ett samband mellan förekomst av hoppkräftor och kolerabakterier i orenat vatten, eftersom bakterierna fäster sig på ytan av djur i plankton. Larven till den besvärliga parasitiska masken Dracunculus (guineamask) måste också utvecklas i copepoder innan den överförs till människa. Infektionsrisken minskar betydligt om vattnet filtreras genom ett enkelt filter av textilmaterial.
Hoppkräftor har använts framgångsrikt i Vietnam för att bekämpa sjukdomsspridande myggor som t.ex. Aedes aegypti, som överför denguefeber och andra sjukdomar. Copepoderna tillsätts till vattenreservoarer där myggorna förökar sig. De (särskilt släktena Mesocyclops och Macrocyclops) kan överleva i flera månader i reservoarerna. De angriper, dödar och äter upp mygglarverna i dessas första två stadier efter kläckningen. Denna biologiska bekämpning kompletteras med avlägsnande av avfall m.m. för att minska myggens yngelplatser. Eftersom vattnet i dessa reservoarer kommer från regnvatten och inte är förorenat är risken att tillföra kolerabakterier liten, och inget fall av kolera har kunnat kopplas till tillsatta copepoder i vattenreservoarer. Metoden prövas även i Thailand och södra USA.[6][7]
Hoppkräftor (Copepoda) är små kräftdjur, några mm i längd, som förekommer i både hav och sötvatten. En del arter är plankton (frisimmande), en del är bentiska (bottenlevande). De lever som filtrerare, rovdjur eller parasiter. Med 13 000 arter är de den artrikaste gruppen bland kräftdjuren och utgör en huvuddel av marint zooplankton. Dessa skenbart obetydliga kräftdjur är kanske störst i djurriket både beträffande individantal och biomassa och en ytterst viktig del av de marina näringskedjorna. De har avgörande betydelse för bestånden av fisk och högre djur. De har också mycket stor betydelse för kolets kretslopp och därmed klimatet.
Några kända hoppkräftor är de mm-stora Cyclops som är mycket vanliga i sötvatten, de större Calanus som dominerar i kalla hav och Harpacticider som är små och bottenlevande i haven.
Arthropoda Şubesine ait Crustacea altşubesi içerisinde yer alan Maxillopoda sınıfının altı altsınıfından biri olan Copepoda, ismini yüzme bacaklarının morfolojik özelliklerine dayanarak Yunanca “kope” (kürek) ve “podos” (ayak) kelimelerinin birleşiminden almıştır. Kopepodlar oldukça eski evrimsel tarihe sahip olan eklembacaklılardır. Vücutlarının oldukça küçük ve dayanıksız olmaları nedeniyle fosilleşmeleri oldukça zordur.[1] Güney İngiltere’deki neolitik kazı alanında harpaktikoid bir kopepod olan Enhydrosoma garienis ’in bir erkek bireyi tanımlanmıştır; bu birey fosilden daha çok kurumuş durumda olup rehidrasyon işlemi ile incelenebilir duruma getirilip teşhis edilmiitir. Kopepodlara ait ilk gerçek fosil kayıtları Palmer tarafından Kuzey ve Güney Amerika’da Miyosen ve Pleistosen döneminden kalma bor mineralleri içerisinde bulunan bireylerden elde edilmiştir[2] .Bu bireylerden bir tanesi Cletocamptus Schmankewitsch,cinsi olarak tanımlanırken diğeri takım seviyesine kadar teşhis edilebilmiştir.En dikkat çekici fosil kopepod hiç şüphesiz Kabatarina pattersoni dir. Brezilya’da Alt Kretase’ye ait kalker kayaçlarda bulunan bu tür parazitik olup yine fosil tür olan Cladocyclus gardneri ,solungaçlarında tespit edilmiştir.[3] Bu veriler kopepodlar'ın fosil kayıtlarının günümüzden 110-120 MY öncesine kadar geri çekilmesini sağlamıştır.Çok uzun evrimsel tarihleri boyunca kopepodlar denizler, yeraltı suları, sürekli ve geçici tatlı ve acı sular ile nemli karasal habitatları da içeren tüm sucul biyotoplarda başarılı bir şekilde kolonize olmuşlardır. Antarktik göllerin donmuş yüzeyleri altında ve Nepal Dağları’nda 5540 m yükseklikte dahi bulunmuşlardır.[4] Bununla birlikte kopepodların yaşam alanları içine tatlısu ve denizel sedimentler, yeraltı suları, bataklıkların yanı sıra ağaç kovuklarındaki su birikintileri ve hatta atık araba tekerlekleri ve bromeliadların yaprakları arasında biriken su ortamları gibi kriptik habitatlar da bulunmaktadır. Ayrıca, serbest yaşayan siklopoid ve harpaktikoid kopepodlar orman tabanındaki nemli çürümüş bölge faunasının da önemli bileşenleridir.[5] Derin hidrotermal kaynaklardan tespit edilen omurgasızların en baskın ve zengin grubunu da kopepodlar oluşturmaktadır[5] Sadece bu ortamda yaşayan bir familya olan Dirivultidae 13 cinse dahil 52 tür içermektedir.[5] Kopepodların günümüzde 10000’e yakın türü bilinmektedir fakat çalışmaların devamı ile birlikte Siphonostomatoida, Poecilostomatoida ve Harpacticoida gibi takımların gerçek çeşitlilikleri ortaya çıkarıldıkça tür sayısının bulunduğumuz yüzyıl ortasına kadar iki katına çıkabileceği tahmin edilmektedir.[6]
Arthropoda Şubesine ait Crustacea altşubesi içerisinde yer alan Maxillopoda sınıfının altı altsınıfından biri olan Copepoda, ismini yüzme bacaklarının morfolojik özelliklerine dayanarak Yunanca “kope” (kürek) ve “podos” (ayak) kelimelerinin birleşiminden almıştır. Kopepodlar oldukça eski evrimsel tarihe sahip olan eklembacaklılardır. Vücutlarının oldukça küçük ve dayanıksız olmaları nedeniyle fosilleşmeleri oldukça zordur. Güney İngiltere’deki neolitik kazı alanında harpaktikoid bir kopepod olan Enhydrosoma garienis ’in bir erkek bireyi tanımlanmıştır; bu birey fosilden daha çok kurumuş durumda olup rehidrasyon işlemi ile incelenebilir duruma getirilip teşhis edilmiitir. Kopepodlara ait ilk gerçek fosil kayıtları Palmer tarafından Kuzey ve Güney Amerika’da Miyosen ve Pleistosen döneminden kalma bor mineralleri içerisinde bulunan bireylerden elde edilmiştir .Bu bireylerden bir tanesi Cletocamptus Schmankewitsch,cinsi olarak tanımlanırken diğeri takım seviyesine kadar teşhis edilebilmiştir.En dikkat çekici fosil kopepod hiç şüphesiz Kabatarina pattersoni dir. Brezilya’da Alt Kretase’ye ait kalker kayaçlarda bulunan bu tür parazitik olup yine fosil tür olan Cladocyclus gardneri ,solungaçlarında tespit edilmiştir. Bu veriler kopepodlar'ın fosil kayıtlarının günümüzden 110-120 MY öncesine kadar geri çekilmesini sağlamıştır.Çok uzun evrimsel tarihleri boyunca kopepodlar denizler, yeraltı suları, sürekli ve geçici tatlı ve acı sular ile nemli karasal habitatları da içeren tüm sucul biyotoplarda başarılı bir şekilde kolonize olmuşlardır. Antarktik göllerin donmuş yüzeyleri altında ve Nepal Dağları’nda 5540 m yükseklikte dahi bulunmuşlardır. Bununla birlikte kopepodların yaşam alanları içine tatlısu ve denizel sedimentler, yeraltı suları, bataklıkların yanı sıra ağaç kovuklarındaki su birikintileri ve hatta atık araba tekerlekleri ve bromeliadların yaprakları arasında biriken su ortamları gibi kriptik habitatlar da bulunmaktadır. Ayrıca, serbest yaşayan siklopoid ve harpaktikoid kopepodlar orman tabanındaki nemli çürümüş bölge faunasının da önemli bileşenleridir. Derin hidrotermal kaynaklardan tespit edilen omurgasızların en baskın ve zengin grubunu da kopepodlar oluşturmaktadır Sadece bu ortamda yaşayan bir familya olan Dirivultidae 13 cinse dahil 52 tür içermektedir. Kopepodların günümüzde 10000’e yakın türü bilinmektedir fakat çalışmaların devamı ile birlikte Siphonostomatoida, Poecilostomatoida ve Harpacticoida gibi takımların gerçek çeşitlilikleri ortaya çıkarıldıkça tür sayısının bulunduğumuz yüzyıl ortasına kadar iki katına çıkabileceği tahmin edilmektedir.
Веслоно́гі ра́ки (Copepoda) — підклас водних ракоподібних класу Щелепоногих (Maxillopoda). Незважаючи на те, що ці створіння надзвичайно малі, вони поширені в морі у величезних кількостях. Веслоногі належать до групи зоопланктону. Вони збирають свою їжу з води за допомогою дрібнопористих волосинок, що знаходяться на ділянках рота. Також, ці раки їдять тварин і водоростей менших, ніж вони самі. Багато морських тварин, наприклад, молюски, асцидії, і такi риби, як оселедець і скумбрія і навіть різні види китів, живуть за рахунок цих міні-ракоподібних. Це робить Веслоногих раків найважливішою ланкою між мікроскопічним планктоном та іншими тваринами які живуть у воді.
Веслоногі раки підклас переважно морських тварин який належать до підтипу ракоподібних (ракоподібні). Веслоногі раки діляться на десять рядів. Налічує 13 000 видів веслоногих раків, та як відомо, 2800 з них живуть в прісній водоймах.
Веслоно́гі ра́ки (Copepoda) — підклас водних ракоподібних класу Щелепоногих (Maxillopoda). Незважаючи на те, що ці створіння надзвичайно малі, вони поширені в морі у величезних кількостях. Веслоногі належать до групи зоопланктону. Вони збирають свою їжу з води за допомогою дрібнопористих волосинок, що знаходяться на ділянках рота. Також, ці раки їдять тварин і водоростей менших, ніж вони самі. Багато морських тварин, наприклад, молюски, асцидії, і такi риби, як оселедець і скумбрія і навіть різні види китів, живуть за рахунок цих міні-ракоподібних. Це робить Веслоногих раків найважливішою ланкою між мікроскопічним планктоном та іншими тваринами які живуть у воді.
Размеры большинства видов составляют от 1 до 10 мм, есть более мелкие интерстициальные виды и более крупные (до 2—3 см) глубоководные и паразитические. Паразитические копеподы часто достигают намного более внушительных размеров, так например Kroyeria caseyi достигает 6,5 см[4].
Тело копепод делится на три тагмы: голову — цефалосому (в копеподологии иногда её называют цефалоторакс, головогрудь), грудь (торакс) и брюшко (абдомен). При этом тельсон (анальную лопасть) многие копеподологи называют последним брюшным (анальным) сегментом.[5]
Тело копепод может «складываться» пополам, изгибаясь в сагиттальной плоскости. При этом граница между функционально передним отделом тела (просомой) и функционально задним (уросомой) у циклопоид и гарпактицид проходит между сегментами груди, несущими четвёртую и пятую пары ног. Эти группы объединяют под названием «Podoplea» — «ногобрюхие». У каланоид граница между просомой и уросомой проходит позади сегмента, несущего пятую пару ног, поэтому их называют «Gymnoplea» — «голобрюхие». Этому признаку, который хорошо коррелирует с другими особенностями строения, придаётся высокий таксономический вес, а Podoplea и Gymnoplea рассматриваются как таксономические категории (в современных классификациях копепод — как надотряды).
Голова несёт одноветвистые антенны первые (антеннулы, усы), двуветвистые антенны вторые (антенны), мандибулы, максиллы первые (максиллулы), максиллы вторые (максиллы) и максиллипеды (ногочелюсти) — придатки сросшегося с головой первого грудного сегмента. У представителей большинства семейств гарпактицид и у некоторых представителей других отрядов с головой сливается и следующий сегмент груди, несущий плавательные конечности, которые при этом могут сильно видоизменяться.
На голове, между мандибулами, располагается ротовое отверстие, прикрытое спереди крупной верхней губой, а сзади — небольшой нижней губой. На переднем крае головы находится направленный вниз вырост — рострум, иногда отчленённый. Антенны I (антеннулы) всегда одноветвистые. Число их члеников различается у представителей разных отрядов. Так, у гарпактицид обычно 5—8 члеников (у самцов до 14); у большинства каланоид 21—27 члеников; у циклопоид от 9 до 23 члеников. У типичных представителей различается относительная длина антеннул: у каланоид они примерно равны телу, у циклопоид равны головогруди, а у гарпактицид заметно короче головогруди. Антенны I участвуют в локомоции, а также несут сенсиллы.
Антенны II обычно двуветвистые (у многих циклопоид одноветвистые), участвуют в создании токов воды для плавания и питания. Мандибула подразделяется на коксу, которая образует жевательный вырост (гнатобазу) с зубцами и щетинками, и щупик, который исходно состоит из базиса, экзо- и эндоподита. Часто ветви, а иногда и основание щупика редуцируются. Так, у многих циклопов от мандибулы отходят лишь три щетинки, которые считаются рудиментом щупика. На жевательных зубцах мандибулы многих морских копепод есть «коронки» из оксида кремния, которые помогают им разгрызать прочные домики диатомовых водорослей.
На следующих за ногочелюстным сегментом четырёх сегментах груди располагаются двуветвистые плавательные конечности — ноги уплощённой формы, служащие основными двигателями при плавании, за наличие которых отряд и получил своё название. Плавательная конечность состоит из двучленикового протоподита, базальный (проксимальный) членик которого называется кокса, а дистальный — базис, и двух отходящих от базиса ветвей (иногда считается, что в состав протоподита входит ещё один членик — прекокса, который слабо отчленён от тела). Наружная (экзоподит) и внутренняя (эндоподит) ветви состоят из 2—4 члеников каждая и несут длинные щетинки, покрытые длинными тонкими выростами (сетулами), и более короткие шипы.
На последнем сегменте груди расположена пятая пара грудных ног, которые обычно не участвуют в плавании и у многих групп сильно уменьшены или видоизменены. У самцов большинства семейств каланоид они резко асимметричны. Эндоподиты обеих ног часто рудиментарны, экзоподит одной из ног служит для переноса сперматофора к семяприёмникам самки при спаривании, а более крупный экзоподит другой конечности несёт длинный когтевидно изогнутый шип, участвующий в удержании самки. Строение и вооружение пятой пары ног для циклопоид и каланоид служит важнейшим таксономическим признаком.
Брюшко состоит обычно из 2—4 сегментов (не считая тельсона). На первом сегменте брюшка находятся парные половые отверстия. У гарпактицид и циклопоид на нём расположена рудиментарная шестая пара ног, у каланоид он лишён конечностей. Остальные сегменты брюшка конечностей не несут. На тельсоне располагаются два подвижных придатка — вилочка, или фурка (фуркальные ветви). Эти придатки состоят из одного членика и не гомологичны конечностям. На фурке находятся фуркальные щетинки, длина и расположение которых — важный таксономический признак.
Как правило, у самок сливаются первый и второй сегменты брюшка, образуя крупный генитальный сегмент; у самцов такого слияния не происходит, поэтому у самцов на один брюшной сегмент больше, чем у самок. У представителей Cyclopoida и Harpacticoida самцы обычно заметно мельче самок, имеют крючковидно изогнутые, укороченные антенны I, служащие для схватывания и удержания самок при спаривании. У многих Calanoida самки и самцы по размерам не различаются. У самцов имеется одна видоизменённая антенна I, которая называется геникулирующая антенна. Она расширена в средней части и способна «складываться пополам»; как и у циклопов, она служит для удержания самки при спаривании. В отдельных случаях наблюдается половой диморфизм в строении практически любых пар конечностей и сегментов тела[6].
Центральная нервная система состоит из головного мозга и соединённой с ним окологлоточным нервным кольцом брюшной нервной цепочки. От головного мозга отходит непарный нерв к науплиальному глазу и парные нервы к фронтальному органу, а также нервы к антеннулам и антеннам (последние — от тритоцеребрума). В состав подглоточного ганглия входят ганглии мандибул, первых и вторых максилл. Ганглии брюшной нервной цепочки слабо отграничены друг от друга. Вся брюшная нервная цепочка расположена в головогруди, в брюшко она не заходит [7].
У многих видов копепод аксоны окружены миелиновой оболочкой [8][9], что встречается редко среди беспозвоночных (другие примеры — некоторые аннелиды и высшие ракообразные, такие как креветки-палемониды и пенеиды). Более того, у копепод миелин высоко организован и напоминает своей многослойной структурой таковой челюстноротых позвоночных.
Узкий эктодермальный пищевод направлен от ротового отверстия к спинной стороне тела и назад. Он переходит в расширенную энтодермальную среднюю кишку, которая, постепенно сужаясь, тянется через всю головогрудь и большую часть брюшка. В задней части брюшка средняя кишка переходит в короткую эктодермальную заднюю кишку, которая открывается анальным отверстием на заднем крае тельсона, между основаниями фуркальных ветвей.
У каланоид в первом свободном грудном сегменте над кишечником расположено яйцевидное сердце, снабжённое одной парой остий — отверстий, через которые в него попадает гемолимфа. Кровеносных сосудов нет, через отверстие в передней части сердца гемолимфа изливается в полость тела. У представителей большинства других отрядов копепод отсутствует и сердце, так что (редкий случай среди членистоногих) кровеносная система полностью редуцирована. При этом у циклопоид гемолимфа (полостная жидкость) перебалтывается в полости тела за счёт движений кишечника, который поднимают и опускают специальные мышцы.
У копепод полностью отсутствуют специальные органы дыхания, что довольно необычно для членистоногих. Мелкие размеры и обилие выростов, увеличивающие относительную площадь поверхности, а также очень небольшая толщина хитинового покрова позволяют копеподам дышать всей поверхностью тела[источник не указан 33 дня].
На личиночных стадиях органами выделения служат парные антеннальные (расположенные в сегменте антенн вторых) железы, а на взрослой стадии — максиллярные (расположенные в сегменте вторых максилл). Считается, что и те, и другие представляют собой видоизменённые целомодукты. Они состоят из замкнутого мешочка (остатка целомической полости) и извитого выводного канала.
Многие копеподы — типичные представители зоопланктона, имеющие характерные адаптации к жизни в толще воды. Это прежде всего приспособления, позволяющие удерживаться в толще воды с минимальными затратами энергии: мелкие размеры (за счёт этого увеличивается относительная площадь поверхности и отношение величины силы трения к массе тела); наличие увеличивающих площадь трения о воду выростов (антеннулы, щетинки антеннул и фурки); тонкие покровы; запасание жира, уменьшающего плотность тела.
Большинство свободноживущих копепод питаются одноклеточными или мелкими колониальными водорослями, которых они отфильтровывают в толще воды, а также донными диатомовыми, бактериями и детритом, которые они могут собирать или соскабливать на дне. Многие виды каланоид и циклопоид — хищники, поедающие другие виды ракообразных (молодь копепод и кладоцер), коловраток, личинок насекомых I—II возраста (в том числе личинок хирономид и кулицид) и др. Копеподитные стадии некоторых пресноводных циклопоид залезают в выводковые камеры дафний, где пожирают яйца[10].
При более детальном изучении «фильтрационного» питания копепод с помощью скоростной микрокиносъёмки выяснилось, что многие из них «охотятся» на отдельные клетки водорослей, которые отлавливают поодиночке. Питающиеся водорослями копеподы запасают энергию пищи в жировых каплях, которые содержатся в их тканях и часто окрашены в желтовато-оранжевый цвет. У полярных видов, питающихся в первую очередь диатомовыми, в период массового весеннего «цветения» объём жировых запасов может достигать половины объёма тела.
Большинство видов копепод — раздельнополые животные. Практически всем им свойственен половой диморфизм, особенно ярко выраженный у многих паразитических видов.
Спариванию предшествует сложное половое поведение, в котором, видимо, обычно играют важную роль как хеморецепция, так и механорецепция. Самки копепод выделяют половые феромоны, которые воспринимаются самцами при помощи хемосенсорных щетинок (эстетасков) первых антенн [11].
При спаривании у большинства семейств каланоид самец сперва захватывает самку за тельсон или фуркальные ветви при помощи геникулирующей антенны, затем — за участок тела, расположенный перед или сразу после генитального сегмента при помощи ног пятой пары, при этом самец и самка обычно располагаются «головой к хвосту» друг друга[12]. Спаривание продолжается от нескольких минут до нескольких суток.
Свободноживущие копеподы имеют сперматофорное оплодотворение. Крупные, сравнимые по размерам с размерами брюшка животного, сперматофоры каланоид переносятся на генитальный сегмент самки во время спаривания при помощи левой пятой ноги самца; на её конце имеются «щипчики», удерживающие бутылковидный сперматофор за суженную базальную часть.
Из яиц копепод выходит личинка ортонауплиус, имеющая три передних пары конечностей — одноветвистые антенны I и двуветвистые антенны II и мандибулы. После первой линьки ортонауплиус превращается в метанауплиуса, у которого имеются зачатки следующей пары конечностей (максилл II).
Светятся представители многих семейств каланоид, а также Oncaea conifera (Poecilostomatoida) [1]. У последней светящийся секрет не выделяется наружу, как это обычно происходит у других копепод, но остаётся внутри многочисленных одноклеточных желёз, сообщающихся порами с внешней средой. Эти железы выдают короткие (80—200 мс) вспышки, до 30 вспышек в секунду и до нескольких сотен вспышек подряд до истощения люминесценции.
У всех изученных в этом отношении копепод люциферином служит широко распространённый среди морских организмов целентеразин [2].
Копеподы играют исключительно важную роль в водных экосистемах и во всей биосфере. Видимо, они имеют наибольшую биомассу среди всех групп водных животных и почти наверняка занимают первое место по доле во вторичной продукции водоёмов. Как потребители фитопланктона копеподы являются главными консументами I порядка в морях и пресных водах. Копеподы служат основной пищей для многих других водных животных, от стрекающих и гребневиков до усатых китов.
Поверхностные воды океанов в настоящее время считаются самым крупным поглотителем углекислого газа (2 миллиарда тонн в год — вероятно, около трети антропогенной эмиссии). Во многом поглощение избыточной углекислоты обеспечивается жизнедеятельностью копепод.
Многие планктонные копеподы питаются ночью в поверхностных слоях океана, а днём мигрируют на глубину, чтобы избежать выедания зрительными хищниками. Их трупы, сброшенные экзувии и фекальные пеллеты опускаются в более глубокие слои воды. Это (как и дыхание живых копепод,переваривающих пищу на глубине) способствует перемещению биогенного углерода из верхних слоёв воды в донные осадки. Кроме того, образование в ходе питания копепод фекальных пеллетов способствует очистке верхних слоёв воды от минеральных взвесей. Это увеличивает прозрачность воды и, следовательно, продукцию фитопланктона.
По разным оценкам, от трети до половины всех видов копепод — комменсалы и паразиты разных групп животных.
Хозяевами копепод могут быть сифонофоры, коралловые полипы, моллюски, кольчатые черви, ракообразные, иглокожие, оболочники, а также костные и хрящевые рыбы, амфибии и водные млекопитающие[13].
Благодаря своей распространённости и широком представительстве среди планктона, копеподы обеспечивают питание многих рыб, тем самым являясь для человека полезными; в то же время, паразитические копеподы поголовье рыб снижают, но это входит в механизм естественного регулирования численности последних[источник не указан 1942 дня]. Представители копеподов циклопы являются промежуточными хозяевами опасного паразита человека – широкого лентеца.
В одной из недавних сводок по систематике ракообразных (Martin, Davis, 2001) предлагается рассматривать копепод в качестве подкласса, который делится на два инфракласса и несколько отрядов:
В предложенной системе копеподы делятся на 218 семейств.
В 2017 году система была значительно переработана, отряд Harpacticoida распался на 2 отряда, а Poecilostomatoida и Cyclopoida стали одним отрядом[14]:
CopepodaCyclopoida (+ Poecilostomatoida)
Тело копепод делится на три тагмы: голову — цефалосому (в копеподологии иногда её называют цефалоторакс, головогрудь), грудь (торакс) и брюшко (абдомен). При этом тельсон (анальную лопасть) многие копеподологи называют последним брюшным (анальным) сегментом.
Тело копепод может «складываться» пополам, изгибаясь в сагиттальной плоскости. При этом граница между функционально передним отделом тела (просомой) и функционально задним (уросомой) у циклопоид и гарпактицид проходит между сегментами груди, несущими четвёртую и пятую пары ног. Эти группы объединяют под названием «Podoplea» — «ногобрюхие». У каланоид граница между просомой и уросомой проходит позади сегмента, несущего пятую пару ног, поэтому их называют «Gymnoplea» — «голобрюхие». Этому признаку, который хорошо коррелирует с другими особенностями строения, придаётся высокий таксономический вес, а Podoplea и Gymnoplea рассматриваются как таксономические категории (в современных классификациях копепод — как надотряды).
Голова и её придаткиГолова несёт одноветвистые антенны первые (антеннулы, усы), двуветвистые антенны вторые (антенны), мандибулы, максиллы первые (максиллулы), максиллы вторые (максиллы) и максиллипеды (ногочелюсти) — придатки сросшегося с головой первого грудного сегмента. У представителей большинства семейств гарпактицид и у некоторых представителей других отрядов с головой сливается и следующий сегмент груди, несущий плавательные конечности, которые при этом могут сильно видоизменяться.
На голове, между мандибулами, располагается ротовое отверстие, прикрытое спереди крупной верхней губой, а сзади — небольшой нижней губой. На переднем крае головы находится направленный вниз вырост — рострум, иногда отчленённый. Антенны I (антеннулы) всегда одноветвистые. Число их члеников различается у представителей разных отрядов. Так, у гарпактицид обычно 5—8 члеников (у самцов до 14); у большинства каланоид 21—27 члеников; у циклопоид от 9 до 23 члеников. У типичных представителей различается относительная длина антеннул: у каланоид они примерно равны телу, у циклопоид равны головогруди, а у гарпактицид заметно короче головогруди. Антенны I участвуют в локомоции, а также несут сенсиллы.
Антенны II обычно двуветвистые (у многих циклопоид одноветвистые), участвуют в создании токов воды для плавания и питания. Мандибула подразделяется на коксу, которая образует жевательный вырост (гнатобазу) с зубцами и щетинками, и щупик, который исходно состоит из базиса, экзо- и эндоподита. Часто ветви, а иногда и основание щупика редуцируются. Так, у многих циклопов от мандибулы отходят лишь три щетинки, которые считаются рудиментом щупика. На жевательных зубцах мандибулы многих морских копепод есть «коронки» из оксида кремния, которые помогают им разгрызать прочные домики диатомовых водорослей.
Грудь и её придаткиНа следующих за ногочелюстным сегментом четырёх сегментах груди располагаются двуветвистые плавательные конечности — ноги уплощённой формы, служащие основными двигателями при плавании, за наличие которых отряд и получил своё название. Плавательная конечность состоит из двучленикового протоподита, базальный (проксимальный) членик которого называется кокса, а дистальный — базис, и двух отходящих от базиса ветвей (иногда считается, что в состав протоподита входит ещё один членик — прекокса, который слабо отчленён от тела). Наружная (экзоподит) и внутренняя (эндоподит) ветви состоят из 2—4 члеников каждая и несут длинные щетинки, покрытые длинными тонкими выростами (сетулами), и более короткие шипы.
На последнем сегменте груди расположена пятая пара грудных ног, которые обычно не участвуют в плавании и у многих групп сильно уменьшены или видоизменены. У самцов большинства семейств каланоид они резко асимметричны. Эндоподиты обеих ног часто рудиментарны, экзоподит одной из ног служит для переноса сперматофора к семяприёмникам самки при спаривании, а более крупный экзоподит другой конечности несёт длинный когтевидно изогнутый шип, участвующий в удержании самки. Строение и вооружение пятой пары ног для циклопоид и каланоид служит важнейшим таксономическим признаком.
Брюшко и его придаткиБрюшко состоит обычно из 2—4 сегментов (не считая тельсона). На первом сегменте брюшка находятся парные половые отверстия. У гарпактицид и циклопоид на нём расположена рудиментарная шестая пара ног, у каланоид он лишён конечностей. Остальные сегменты брюшка конечностей не несут. На тельсоне располагаются два подвижных придатка — вилочка, или фурка (фуркальные ветви). Эти придатки состоят из одного членика и не гомологичны конечностям. На фурке находятся фуркальные щетинки, длина и расположение которых — важный таксономический признак.
Половой диморфизмКак правило, у самок сливаются первый и второй сегменты брюшка, образуя крупный генитальный сегмент; у самцов такого слияния не происходит, поэтому у самцов на один брюшной сегмент больше, чем у самок. У представителей Cyclopoida и Harpacticoida самцы обычно заметно мельче самок, имеют крючковидно изогнутые, укороченные антенны I, служащие для схватывания и удержания самок при спаривании. У многих Calanoida самки и самцы по размерам не различаются. У самцов имеется одна видоизменённая антенна I, которая называется геникулирующая антенна. Она расширена в средней части и способна «складываться пополам»; как и у циклопов, она служит для удержания самки при спаривании. В отдельных случаях наблюдается половой диморфизм в строении практически любых пар конечностей и сегментов тела.
橈腳類(學名:Copepods;/ˈkoʊpɪpɒd/,「槳足」的意思)是一類細小的甲殼類動物,生活在海洋及差不多所有淡水的棲地,亦是海洋中重要的蛋白質來源 [1]。很多的橈腳類都是浮游動物,當中有些是底棲生物,一些則是湖沼陸棲生物,在陸地上及沼澤、水溝等水體生活。很多都是生活在地底下的,如洞穴、水坑或河床等,甚或雨林內的落葉堆中。橈腳類有時被用作為指標物種(indicator species)。
橈腳類是甲殼亞門下的一個亞綱或是一個綱。牠們之下共有10個目,當中有超過13000個已記述的物種,2800個在淡水中生活[2]。
橈腳類的大小差別很大,但一般長1-2毫米,體形像淚珠,有大的觸角。雖然牠們像其他甲殼類般有裝甲似的外骨骼,不過由於身體太細小,這一層薄薄的裝甲連同包裹着的身軀看起來差不多是完全透明。一些生活在極地的橈腳類可以長達1厘米。橈腳類只有一隻複眼,很多時是鮮紅色及位於頭部的中央。一些在地底的物種甚至連眼睛也沒有。
大部份細小的橈腳類會直接吃浮游植物,很少較大的物種是掠食其他細小的同類。草食性的橈腳類,尤其是生活在冰冷海洋的群族,會將從食物獲得的能量儲存成油滴,這些油滴可以佔了牠們超過一半的身體。
很多橈腳類的神經元有髓磷脂包圍,這在無脊椎動物中是很稀有的。這些髓磷脂很有組織,就像是其他脊椎動物般。
一些橈腳類是很迴避性的,可以極速逃亡幾毫米的距離。
由於橈腳類細小的身形,這些物種基本上不需要心臟或任何循環系統節存在。即使在哲水蚤目(Calanoida)[5]的物種都有心臟,但這些物種的身上沒有血管,也沒有鰓,而是直接把氧氣吸進體內。另一方面,牠們的排洩系統亦只有靠在其口器的一對上頜竇(maxillary glands)。
浮游的橈腳類對全球生態及碳循環有著重要的角色。牠們是浮游動物中優越的成員,且是細小魚類、鯨魚、海鳥及其他甲殼類,如磷蝦的主要食物。一些科學家認為牠們與南極磷蝦形成了地球上最大的生物質量。由於牠們的體型細小,生長率相對較快。加上牠們更為平均分佈在世界各地的海洋,故橈腳類對全球海洋的次級生產及碳匯的貢獻比磷蝦或其他的生物群更多。現時相信海洋表面是世界最大的碳匯,每年吸納了約20億噸的碳,佔人類碳排放的三分之一並減低其影響。很多浮游橈腳類會在夜間到海面覓食,日間回到海底以避開掠食者。牠們脫落的外骨骼、糞便及呼吸作用可以將碳帶到深海。
一些橈腳類是寄生的[7][8],並擁有高度改良的身體。牠們會附在魚類、鯊魚、水中哺乳動物及多類無脊椎動物,如貝類、被囊類或珊瑚。牠們可以是體外或體內寄生的。
橈腳類有時會在供水系統中出現,尤其是沒有過濾的供水[9],例如:紐約市、波士頓、三藩市等地[10]。在已處理的供水系統中並不怎麼有問題,但在如秘魯及孟加拉等熱帶國家,就有發現橈腳類與在未經處理供水中的霍亂有關,而霍亂菌是附在浮游生物的表面。此外,在麥地那龍線蟲的幼蟲可以傳播給人類之前,牠要在橈腳類動物的消化道內才可以發育。 感染這些疾病的風險,可以透過利用布過濾器之類的物品把橈足類及其他物質濾走,來減低問題的影響。
在越南已經成功以橈腳類來控制帶病的蚊,如傳播登革熱的埃及斑蚊[11][12]。將橈腳類(如中劍水蚤及大劍溞屬)加入容器中,牠們可以在容器中生活多個月,攻擊、殺死及掠食孑孓[9]。這類生物控制方法須與清理及回收垃圾等配合,以消除蚊子滋生的地方。
|chapter=被忽略 (帮助) 橈腳類(學名:Copepods;/ˈkoʊpɪpɒd/,「槳足」的意思)是一類細小的甲殼類動物,生活在海洋及差不多所有淡水的棲地,亦是海洋中重要的蛋白質來源 。很多的橈腳類都是浮游動物,當中有些是底棲生物,一些則是湖沼陸棲生物,在陸地上及沼澤、水溝等水體生活。很多都是生活在地底下的,如洞穴、水坑或河床等,甚或雨林內的落葉堆中。橈腳類有時被用作為指標物種(indicator species)。

カイアシ類(橈脚類)とは、節足動物門 甲殻亜門 カイアシ亜綱に属する動物の総称である。現在10目約11,000種が報告されている。ケンミジンコまたは、英名Copepodaのカタカナ読みでコペポーダとも呼ばれる。多くは浮遊生物として生活する微小な甲殻類であるが、底生性、寄生性なども存在する。海洋生態系においては食物連鎖上、重要な位置付けである。
カイアシ(橈脚)類は、甲殻類のカイアシ亜綱に属する動物の総称である。脚が船(ボート)の橈(かい)のような形をしていることから「橈脚(かいあし)」と呼ばれる。英名のCopepod(Cope橈-pod脚)も同様の由来である。カイアシ類の別名が「ケンミジンコ(剣微塵子)」であるが、胴体を剣の刃、第1触角を鍔に見立てた名称である。
大部分は浮遊生活(プランクトン)をしている。他に底生性、間隙性[1]、寄生性[2]もある。一部では地下水性[3][4]や半陸上性[3][5](土壌や落葉、コケなどの水膜中に生息し、完全に陸上で生息しているわけではない;semi-terrestrial)、葉上性[6]も知られる。半陸上性のグループは主に開拓されていない里山の土壌中や落葉中に多くの種が生息するが[7]、例外的に皇居からの発見例もある[8]。
海洋において、プランクトンネットで採集される動物プランクトンで、カイアシ類が数、量ともに最大の割合を占めることが多い。魚やクラゲなどの肉食性動物の餌としてきわめて重要である。食物連鎖では、藻類による光合成で生産されたエネルギーを魚など高次の捕食者へ受け渡すという重要な位置にある。
体形は通常、円筒形~紡錘形。体長は0.3mm~10mmで、多くは1mm程度でとても小さい。詳細な形態を観察するには顕微鏡が必要になる。寄生性のある種(ペンネラ(ヒジキムシ)科 Pennellidae)では体長30cmに達する場合もある。体は大きく分けて頭部と胸部が含まれる前体部と、腹部と尾部が含まれる後体部になり、付属肢の多くは前体部に収まっている。頭部には第1触角、第2触角、上顎、第1下顎、第2下顎、顎脚があり、胸部には第1胸脚~第4あるいは第5胸脚がある。第6胸脚は退化的で、雌では生殖節の生殖孔における生殖蓋の役割がある。腹部には腹肢はなく、尾部に尾肢がある。体節数は頭部1節、胸部6節(第6胸節は「生殖節」)、腹部4節(第4腹節は肛門節)、合計11節(幼生や幼体では減少している;頭部と第1胸節が融合することがある)となるが、雌では第6胸節(生殖節)と第1節が完全あるいは不完全に融合して「生殖複合節」を形成する(原始的な種であるプラティコピア目では分離している)。そのため合計10節になる。ただし、便宜上、生殖複合節の次の節は第1腹節(もとは第2腹節)としている。種によっては生殖複合節に1つまたは2つの卵嚢を付けることがある。他の種は卵塊で水中へ放つ。あるいは、卵をバラバラに産卵する。種により休眠卵(耐久卵)を産卵する。生殖複合節に貯精嚢をもち、精子を貯蔵する事ができる。一度の交尾のみで必要に応じて精子と卵と受精させ、一生涯、産卵することが可能である。
第1触角:感覚器として機能し、多くは触角から複数の刺毛が生えている。振動や水中の化学物質を受容する。カラヌス目では前体部程度の長さがある。カラヌス目セントロパジェス科(Centropagidae)では雄で片側の触角が変形し、交尾時に雌を捕まえるための把握器になっている。カラヌス目以外の種では触角は短く、欠くものもいる。
第2触角:第1触角と比べると著しく小型化しており解剖をしないと観察することはできない。遊泳や摂食時の餌をかき集める役割がある。寄生性のあるグループでは宿主へ付着するために鉤爪状へ変形したものもいる。
上顎:餌を咀嚼する器官で、複数の咀嚼歯をもつ。種によっては成体になると退化して摂食をしなくなるものもいる。
第1下顎:餌を捕まえる把握器として機能がある。底節と基節には刺毛が生えており、遊泳としての役割も持つ。
第2下顎:餌を把握する。第1下顎と合わせて餌粒子を濾しとる、懸濁物摂食(suspension feeding)の要になり、これらが位置する部分をフィーディング・バスケット(feeding basket)と称することがある。
顎脚:把握や遊泳としての役割を持つ
胸脚:4対あるいは5対、稀に6対ある。多くは5対で、通常は遊泳として機能する。ハルパクチクス目(ソコミジンコ目;Harpacticoida)では歩脚に変形したものや、第1胸脚が把握としての役割をもつものもいる。カラヌス目では雄の第5胸脚が交尾時に雌を把握し、雌の生殖孔へ精包(spermatophore)をつける働きをもつ。第6胸脚は雄で欠損し、雌では小型化し生殖孔の生殖蓋に変形している。
ノープリウス眼(nauplius eye)と呼ばれる眼は第一触角の間にあり、一つ目の単眼である。代表的な属のひとつ、キクロプス(Cyclops)属の属名はギリシャ神話の一つ目巨人キュクロープスに由来する。種によっては2つ目や5つ目、または眼を欠くものもいる。視力は明度を認識する程度しかない。しかし、ポンテラ科(Pontellidae)やコリケウス科(Corycaeidae)などのグループでは眼が発達し大きなレンズ眼となり[9][10][11]、形態を認識する視力を持っているとされている[9]。
カラヌス目(Calanoida)とミソフリア目(Misophrioida)の一部は心臓をもち[12]、第1胸節または第2胸節の背面に位置する。側面に2つ、後方に1つの開口があり、前方に大動脈がある。ただし大動脈を欠く種もいる。心拍数は1分間に300回程度ととてもはやい。心臓を出た血液は体腔内の小溝中を循環する。血色素はヘムエリスリンだが、甲殻類で一般的なヘモシアニンは報告されていない[13]。
卵から孵化すると、ノープリウス幼生、コペポディット幼体を経て成体になる。ノープリウス幼生はノープリウス第1~6期、コペポディット幼生はコペポディット第1~5期、成体はコペポディット第6期に相当する。計12つのステージがある。寄生性のグループではステージ数を短縮することが知られている[2]。それぞれのステージは脱皮をすることで次のステージに移行し、成体になると脱皮をしなくなる。したがって、一生の脱皮回数は11回となる。
ハルパクチクス目(Harpscticoida)の一部とキクロプス目(Cyclopoida)の一部、カラヌス目(Calanoida)の一部のグループは生息に不敵な状況の間(例えば、低酸素、高温、干ばつ)休眠する。コペポディット幼体や稀に成体のときに休眠するが、中には休眠卵(耐久卵)[14]やシスト(粘液物を分泌して体を包み込みカプセル状に硬化)[15][16]になるものもいる。
コペポディット幼体または成体の休眠は体内に多くの油球(oil sac)を形成する。この油球体積比(いわば体脂肪率)は60%を越える。脂質の種類は多くはワックスエステルとトリグリセリドで[17]、油球中にはワックスエステルが含まれ、トリグリセリドは体内中に分散している[18]。一部にプリステイン(pristane)があり、中性浮力として機能していると考えられている[19]。トリグリセリドをもつ個体は休眠以外でも見られ、休眠個体にはワックスエステルを多く貯蔵している[20]。ワックスエステルはエネルギー貯蔵として働くが、トリグリセリドは海水よりも比重が小さく、浮力としての役割があると考えられている。こうした油球を多く持った休眠個体は食物連鎖上、重要な栄養源となり[21]、水産物中の脂質成分の分布や動態が大きく関わっている。
休眠卵の形状は海産種で、枝状突起や円盤状構造物を付ける。これに対し、普段の卵(急発卵)は卵殻が平滑で構造物がない。[22]。休眠卵は一定時間を超え、環境条件が好適になると孵化するが、環境条件が好適になるまでは孵化はしない。環境条件が不適なとき、海底には数cmで数万個の休眠卵が堆積している場合がある。300年経過した休眠卵を好適条件にしたところ孵化し、成体まで成長したという例がある[23]。
分類はWorld Register of Marine Species (2017.9.14)[24]に従い、カラヌス目の上科はOhtsuka & Huys (2001)[25]と日本海洋データセンター(JODC)[26]に準じて分類した。和名は図鑑[3][4][27][28][29]を参考にした。
2017年にポエキロストム目(Poecilostomatoida)はキクロプス目(Cyclopoida)内に入り、ハルパクチクス目(Harpacticoida)の一部のグループがカヌエラ目(Canuelloida)に新設された[30]。タウマトプシルス目(Thaumatopsylloida)があったが、シクロプス目(Cyclopoida)内のグループのシノニムであったためタウマトプシルス目は撤回された[24]。現在10目に分類される。
カイアシ類(橈脚類)とは、節足動物門 甲殻亜門 カイアシ亜綱に属する動物の総称である。現在10目約11,000種が報告されている。ケンミジンコまたは、英名Copepodaのカタカナ読みでコペポーダとも呼ばれる。多くは浮遊生物として生活する微小な甲殻類であるが、底生性、寄生性なども存在する。海洋生態系においては食物連鎖上、重要な位置付けである。
요각류(橈脚類, 영어: copepod)는 갑각류에 속하는 아강이다. 서식지는 해수면, 해저면, 늪지 등 다양하다. 서식지 생태계의 건강성을 확인하는 지표로 쓰이기도 한다.
다음은 범절지동물의 계통 분류이다.[1]
범절지동물 Tactopoda 진절지동물 대각류 범갑각류탄툴라쿠스류
빈갑류













.jpg)








_A_%26_B_-_Peniculus_hokutae_(Copepoda,_Pennellidae).png)
























.jpg)

