
Browse Trichoplax adhaerens genome and functional genomics using the JGI browser.
Pearse and Voigt (2007) report that in their sampling using glass slides, certain general types of organisms were often found in association with placozoans, including several kinds of sessile ciliates (solitary and colonial vorticellids as well as folliculinids); spirorbid and other serpulid polychaetes; and, in smaller numbers, free-living loxosomatid kamptozoans (entoprocts). Placozoans have been observed capturing and eating ciliates in a culture dish. Wild placozoans are probably opportunistic grazers and scavengers on organic detritus and on algae and bacteria in biofilms covering a diversity of substrates. (Pearse and Voigt 2007 and references therein)
Trichoplax adhaerens, the only described species in the phylum Placozoa, is the structurally simplest metazoan (multicellular animal). It is quite small (just 2 to 3 mm in diameter) and consists of several thousand cells arranged as a double-layered plate. It lacks anterior-posterior polarity and symmetry. However, the cells of the upper and lower layers differ in shape and there is a consistent dorsal-ventral orientation of the body relative to the substrate. Trichoplax moves by ciliary gliding, changing its shape along the edges as it moves, like an amoeba. Very small (presumably young) individuals can swim, but larger individuals crawl. It appears that Trichoplax feeds by phagocytosis of organic detritus. (Brusca and Brusca 2003) Although the discovery and description of Trichoplax adhaerens in 1883 (in saltwater tanks in Austria) sparked controversy among zoologists about what its significance might be with respect to inferring the characteristics of early metazoans, interest faded away as the (incorrect) idea took hold that these enigmatic organisms were simply abnormal larvae of hydrozoan cnidarians. Although strong evidence against this view was published in 1912 and 1914, there was no more discussion of Trichoplax in the zoological literature--nor citation of the key critiques of 1912 and 1914-- for more than half a century (although the misconception that it was a cnidarian was repeated in textbooks). Beginning in the 1960s, several researchers re-focused attention on Trichoplax, demonstrating that it is an adult form of a new phylum dubbed, in 1971, Placozoa. It became clear that placozoans could be found worldwide in the shallow waters of subtropical and tropical regions (Pearse 1989; Eitel and Schierwater 2010). (Syed and Schierwater 2002) Recent data all indicate that the Placozoa represent one of the earliest branching lineages in the metazoan tree, but the exact placement of this branch remains uncertain (Schierwater et al. 2009; Ball and Miller 2010 and references therein).
Pearse and Voigt (2007) summarized all the sites where placozoans were known to have been collected up to the time of their publication. Based on current knowledge, Eitel and Schierwater (2010) predict that most placozoans are found between the equator and 20 degrees North.
Dellaporta et al. (2006) report on the complete mitochondrial genome of Trichoplax adhaerens, which they note is the largest known metazoan mtDNA genome--at 43,079 bp, it is more than twice the size of the typical metazoan mtDNA. Srivastava et al. (2008) present a preliminary analysis of the genome of Trichoplax adhaerens, which is among the smallest known metazoan genomes at about 98 million base pairs.
Placozoans have been found only in shallow, nearshore, tropical and subtropical marine environments between approximately 30 degrees north and south of the Equator. Within these habitats, they tend to be absent from areas of high current and from bare sand bottoms. Because they are very sensitive to reductions in salinity, they will presumably not be found in fresh or brackish water or seasonally in areas that are subjected to heavy monsoon rains. (Ball and Miller 2010 and references therein; Eitel and Schierwater 2010) They are often found on mangrove tree roots, reefs, boat docks in the eulittoral and littoral zone, and around stony beaches (Eitel and Schierwater 2010). They can be abundant on coral reefs. Although placozoans have long been viewed as benthic organisms, Pearse and Voigt (2007) had more success collecting them on glass slides suspended 20 to 60 cm above the bottom than on glass slides situated on the bottom itself.
The placozoan Trichoplax adhaerens is a small (1 to 2 mm) ciliated disc-shaped microscopic marine animal. The thin body consists only of an epithelium and an internal mesenchyme. The epithelium has two regions: an upper free, or dorsal, epithelium of cover cells and a lower attached, or ventral, epithelium of cylinder and gland cells. Some differences between the central and marginal areas in the body have also been reported. When the densely ciliated ventral epithelium is in contact with the substrate, the animals display a gliding or creeping locomotion. When the animal is feeding, its shape changes periodically. (Ueda et al. 1999; Maruyama 2004 and references therein; Srivastava et al. 2008)
When large or stressed, placozoans can undergo binary fission (split into two individuals--sometimes three!) or bud to produce numerous multicellular flagellated "swarmers", which are capable of regenerating the amoeboid stage. However, this is not the complete life cycle, as there is also evidence of sexual reproduction. (Brusca and Brusca 2003; Ball and Miller 2010 and references therein) Sexual reproduction has not been observed in culture, but putative oocyte formation in degenerating animals is seen routinely. Population genetic analyses demonstrate allelic variation and evidence for genetic recombination in animals in the wild that is consistent with sex. (Signorovitch et al. 2005; Srivastava et al. 2008)
Trichoplax adhaerens is the only described species in the phylum Placozoa, but recent work (Eitel and Schierwater 2010 and references therein) suggests that there may be many unrecognized placozoan species.
Lövhəcikkimilər (lat. Placozoa) — Aqnotozoylar yarımaləminə daxil olan heyvan tipi. Bu tipə yalnız çoxhüceyrəli heyvanlar içərisində ən pirimitivi olan Trichoplax adhaerens növü aiddir.
Trichoplax adhaerens és un animal simple amb forma de làmina amb una cavitat corporal plena de fluid pressuritzat. Se li concedí el seu propi fílum, placozous (Placozoa); es creu que hi ha una altra espècie, Treptoplax reptans, descrita l'any 1896 i que no s'ha tornat a trobar, conseqüentment es posa en dubte la seva existència. El nom T. adhaerens li fou donat perquè tendeix a enganxar-se al seu substrat, incloent pipetes de vidre i portaobjectes per a microscopi.
Trichoplax va ser descobert l'any 1883 pel zoòleg alemany Franz Eilhard Schulze, en un aquari d'aigua marina de l'Institut Zoològic a Graz, Àustria. El nom genèric deriva del grec θρίξ (thrix), 'pèl', i πλάξ (plax), 'placa'. L'epítet específic adhaerens prové del llatí, 'adherent', reflectint la tendència a enganxar-se a les làmines de vidre i pipetes utilitzades en el seu examen.[1]
Tot i que des del començament la majoria d'investigadors que estudiaven Trichoplax s'adonaven en qualsevol detall que no tenia cap relació estreta amb cap altre fílum d'animal, el zoòleg Thilo Krumbach va publicar una teoria que proposava que Trichoplax és una forma de la larva plànula de l'hidrozou similar a una anemona Eleutheria krohni (amb el nom acceptat actualment: Eleutheria dichotoma), l'any 1907. Encara que això va ser refutat per Schulze i d'altres, l'anàlisi de Krumbach va esdevenir l'explicació estàndard als llibres de text i res més va ser publicat en les revistes zoològiques sobre Trichoplax fins a la dècada de 1960. En la dècada de 1960 i 1970, un nou interès entre els investigadors va dur a l'acceptació dels Placozoa com a nou fílum animal. Entre les noves descobertes, hi havia l'estudi de les fases primerenques del desenvolupament embrionari d'aquest animal i les proves que aquests animals que s'havien estat estudiant són adults, no larves. Aquest nou interès també incloïa l'estudi de l'organisme en la natura (en comptes dels aquaris).[2]
Un Trichoplax té el cos tou, d'uns 0,5 mm, i pot semblar-se a una ameba. Està constituït per centenars de cèl·lules de quatre tipus diferents en tres capes: cèl·lules monociliades dorsals i epitelials ventrals, cèl·lules glandulars ventrals i fibroses sincitials.
S'ha seqüenciat el genoma mitocondrial complet de Trichoplax adhaerens i els placozous són el fílum més basal de tots els eumetazous vius.[3]
Trichoplax adhaerens és un animal simple amb forma de làmina amb una cavitat corporal plena de fluid pressuritzat. Se li concedí el seu propi fílum, placozous (Placozoa); es creu que hi ha una altra espècie, Treptoplax reptans, descrita l'any 1896 i que no s'ha tornat a trobar, conseqüentment es posa en dubte la seva existència. El nom T. adhaerens li fou donat perquè tendeix a enganxar-se al seu substrat, incloent pipetes de vidre i portaobjectes per a microscopi.
Trichoplax adhaerens ist die einzige allgemein anerkannte Art der Placozoa.[1] Der Organismus wurde 1883 von dem deutschen Zoologen Franz Eilhard Schulze in einem Meerwasser-Aquarium des Zoologischen Instituts in Graz entdeckt. Der von ihm verliehene Gattungsname des Tiers leitet sich aus dem altgriechischen θρίξ thrix „Haar“ und πλάξ plax „Platte“ ab, das lateinische Artepithet adhaerens bedeutet „haftend“, so dass sich der Artenname etwa mit „anhaftende haarige Platte“ übersetzen lässt.
Die Placozoa oder Plattentiere sind die strukturell einfachsten aller vielzelligen Tiere (Metazoa) und bilden einen eigenen Tierstamm. Der wissenschaftliche Name bedeutet wörtlich übersetzt „flache Tiere“, im Deutschen wird gelegentlich der Name Scheibentiere verwendet. Anhand genetischer Daten gilt es als sicher, dass die Placozoa zahlreiche Arten umfassen. Da sie sich aber morphologisch nicht unterscheiden lassen, wurde bisher nur Trichoplax adhaerens als einzige Art beschrieben.[2]
Lange Zeit hielt man die bisher nicht in ihrem natürlichen Lebensraum beobachteten Tiere für Jugendstadien von Nesseltieren (Cnidaria): Länger als ein halbes Jahrhundert wurden sie als Planula-Larven der Hydrozoen-Art Eleutheria krohni gedeutet. Erst Arbeiten aus den 1970er Jahren, unter anderem durch den Tübinger Protozoologen Karl G. Grell, klärten die frühen Phasen der Embryonalentwicklung der Tiere auf und trugen maßgeblich zur Errichtung eines eigenen Tierstamms für sie bei.
Placozoa haben, wie ihr Name andeutet, in der Regel einen stark abgeflachten, scheibenförmigen Körper. Er misst im Durchmesser in den meisten Fällen weniger als einen halben Millimeter. Vereinzelt erreichen Exemplare Größen von 2 bis 3 Millimetern – selten mehr. Die Scheibendicke beträgt meistens nur etwa 25 Mikrometer. Mit bloßem Auge sind die gräulich gefärbten, im Gegenlicht transparenten Tiere meist gerade noch zu erkennen. Oberflächlich sehen sie wie große Amöben aus und ändern wie diese beständig ihre äußere Form. Selten treten daneben kugelförmige Lebensstadien auf, die möglicherweise dem passiven Transport in neue Lebensräume dienen.
Wie die Zuordnung zu den Gewebelosen schon andeutet, fehlen Trichoplax adhaerens Gewebe und Organe; es gibt auch keine ausgeprägte Körpersymmetrie, so dass sich weder vorne oder hinten noch links oder rechts unterscheiden lassen.
Sowohl strukturell als auch funktionell lassen sich bei Trichoplax adhaerens eine Rücken- oder Dorsalseite und eine Bauch- oder Ventralseite unterscheiden. Beide bestehen aus einer einlagigen, nach außen von Schleim bedeckten Zellschicht und erinnern vor allem wegen der zwischen den Zellen bestehenden Verbindungen, der Gürteldesmosomen, an Epithelgewebe. Die Zelllagen der Placozoa besitzen aber im Gegensatz zu einem echten Epithel keine Basallamina; darunter versteht man eine aus extrazellulärem Material bestehende, dem Epithel von innen unterliegende dünne Schicht, die es versteift und vom Innenraum des Körpers abtrennt. Das Fehlen dieser ansonsten bei allen Tieren außer einigen Schwämmen zu findenden Struktur lässt sich funktionell begründen: Eine steife Trennschicht würde die amöboiden Formänderungen von Trichoplax adhaerens unmöglich machen. Um den Unterschied zu betonen, spricht man statt von einem Epithel daher manchmal bei den Placozoa von einem Epitheloid.
Ein ausgewachsenes Individuum besteht aus bis zu tausend Zellen, die sechs verschiedenen Zelltypen[3] zugeordnet werden können (früher waren nur vier bekannt). Die einfach begeißelten Zellen des rückseitigen Epitheloids sind außen abgeflacht und bilden hier einen plattenartigen Zellverband, von dem die Zellkörper ins Innere vorragen, sie enthalten kleine Lipidkörperchen, möglicherweise Abwehrstoffe gegenüber Fressfeinden. Zwischen ihnen befinden sich wenige Zellen mit becherförmigem Zellkern, die einen kristallinen Einschluss enthalten und deshalb Kristallzellen genannt werden; sie könnten eine Funktion als Sinneszellen besitzen. Die Zellen der Bauchseite sind ebenfalls nur einfach begeißelt, sie haben eine langgestreckte säulige Form mit kleinem Querschnitt an der Oberfläche, so dass die Geißeln bauchseitig sehr eng beieinander stehen und eine bewimperte „Kriechsohle“ bilden. Ihre zahlreichen Mikrovilli deuten darauf hin, dass sie auch an der Verdauung von Beute beteiligt sind. Dazwischen, vor allem randlich auf der Ventralseite, befinden sich ebenfalls mit einem Cilium bewimperte Drüsenzellen, die Neuropeptide synthetisieren können. Diese könnten an der Steuerung und Regulation der Bewegung des Tiers beteiligt sein. Ein dritter Typ unbewimperter Zellen, die sogenannten lipophilen Zellen, erstrecken sich mit ihrem Zellkörper weit ins Innere des Organismus, sie sind in der Mitte der Kriechsohle am häufigsten, also genau entgegengesetzt wie die Drüsenzellen verteilt. Vermutlich synthetisieren diese Zellen Verdauungsenzyme.
Zwischen den beiden Zellschichten befindet sich ein flüssigkeitsgefüllter Innenraum, in dem ein lockeres Netzwerk von überwiegend einschichtig angeordneten Faserzellen liegt. Diese bestehen aus dem Zellkörper mit sechs oder mehr langen Fortsätzen, die sich am Ende fein verzweigen. Entgegen früheren Annahmen bilden die Faserzellen wohl kein Synzytium aus.[3] Die Fortsätze ziehen sich zu den Zellen beider Epithelien und zu anderen Faserzellen, sie verknüpfen so alle somatischen Zellen zu einem Netzwerk. Die Kontaktstellen sind morphologisch unspezialisiert, sie bilden keine Synapsen oder ähnliche Bildungen aus. Spärlich in die Faserzellen eingestreute Scheidewände (Septen) könnten aber eine Funktion bei der elektrischen Erregungsleitung besitzen, entsprechende Funktionen sind von ähnlichen Strukturen bei anderen Tierarten bekannt. Stäbchenförmige Einschlüsse könnten endosymbiontische Bakterien unbekannter Funktion sein. Entgegen früheren Annahmen besitzen die Fortsätze keine Myofibrillen, eine Rolle, analog Muskelzellen, bei der Bewegung des Tiers ist damit eher unwahrscheinlich. Ihre Oberfläche trägt nach Immunfluoreszenz-Beobachtungen das Cadherin TaCDH.[3] Es gibt Hinweise darauf, dass unterschiedliche genetische Linien der Placozoa unterschiedliche Faserzellen enthalten.[4]
Soweit bisher bekannt, existiert keine extrazelluläre Matrix wie man sie bei Nesseltieren oder Rippenquallen als Mesogloea oder bei Schwämmen als Mesohyl findet.
Anders als bei den Gewebetieren wurden pluripotente Stammzellen, also solche, die sich in andere Zelltypen umwandeln können, noch nicht eindeutig nachgewiesen.
Alle Zellkerne der Placozoa-Zellen enthalten zwölf nur etwa zwei bis drei Mikrometer große Chromosomen. Drei Paare sind metazentrisch, die anderen acrozentrisch, das heißt, das Zentromer, die Ansatzstelle der Spindelfasern bei der Zellteilung, liegt in der Mitte beziehungsweise am äußeren Rand des jeweiligen Chromosoms.[5] Die Zellen des Fasersynzytiums sind eventuell tetraploid, enthalten also einen vierfachen Chromosomensatz.
Insgesamt enthält ein einfacher Chromosomensatz von Trichoplax adhaerens weniger als 50 Millionen Basenpaare (98 Millionen Basenpaare des diploiden Organismus) und bildet damit das kleinste tierische Genom; die Zahl der Basenpaare des Darmbakteriums Escherichia coli ist nur um einen Faktor 10 kleiner. Es werden zwischen 11.000 und 12.000 proteinkodierende Gene abgeschätzt.[6] Im Gegensatz zu anderen Arten mit kleinem Genom ist dasjenige von Trichoplax nicht an Introns verarmt; das ist ein Indiz gegen eine nachträgliche Verkleinerung.
Die genetische Ausstattung von Trichoplax adhaerens ist noch nicht sehr gut erforscht; einige Gene wie Brachyury oder Tbx2/3, die homolog zu entsprechenden Basenpaarsequenzen bei Gewebetieren sind, konnten allerdings bereits nachgewiesen werden. Von besonderer Bedeutung ist Trox-2, ein Placozoa-Gen, das vermutlich den bei Nesseltieren unter dem Namen Cnox-2 und bei den zweiseitig-symmetrischen Bilateria als Gsx bezeichneten Genen homolog ist, es gehört demnach zu den ParaHox-Genen.[7] Als Homöobox- Gen, das zu den Hox-Genen gehört, spielt es in der Embryonalentwicklung bei Gewebetieren eine Rolle bei der Ausbildung und Körperdifferenzierung entlang der Symmetrieachse; bei den Nesseltieren legt es etwa die Lage von Mund- (oraler) und mundabgewandter (aboraler) Seite des Tieres fest. Placozoa besitzen allerdings, wie angesprochen, keine Symmetrieachsen. Im Trichoplax-Körper wird das Gen in einer ringförmigen, den Körper umgebenden Zone abgelesen. Vergleich der Genanordnung anderer Gene auf dem Chromosomenstrang liefert außerdem Indizien dafür, dass Trichoplax wohl ein weiteres, echtes Hox-Gen besessen haben könnte, das später verloren gegangen ist – ein Indiz für eine sekundäre Vereinfachung.[7]
Das Trichoplax-Genom enthält darüber hinaus zahlreiche weitere Gene, die ortholog zu Genen für Transkriptionsfaktoren sind, die bei Bilateriern (wie auch dem Menschen) an der Ausbildung von Körperachsen beteiligt sind, obwohl diese bei den Placozoa nicht differenziert sind. So sind Gene für den Wnt-Signalweg und den BMP/TGF beta Signalweg vorhanden, der bei Bilateria an der Ausbildung der Körperlängsachse und der Dorsal-Ventral-Achse beteiligt ist. Obwohl spezialisierte Nervenzellen nicht ausgebildet sind, sind Proteine für Neurotransmitter und für Nervenzellen typische Ionenkanäle vorhanden. Wie typisch für Mehrzeller (aber auch schon in vielen einzelligen oder Zellkolonien bildenden Kragengeißeltierchen vorhanden) werden Gene für zahlreiche Zelladhäsionsmoleküle gefunden, darunter auch solche, die an Moleküle der extrazellulären Matrix binden, obwohl eine solche in Trichoplax noch nicht sicher nachgewiesen wurde – möglicherweise ist diese doch vorhanden, aber den bisherigen Nachweismethoden entgangen.[6]
Genaue Verbreitungsangaben existieren für die Placozoa nicht, die Tiere wurden aber unter anderem im Roten Meer, dem Mittelmeer, der Karibik, vor Hawaii, Guam, Samoa, Japan, Vietnam oder Papua-Neuguinea und am Großen Barriere-Riff vor der australischen Ostküste nachgewiesen. Sie sind, soweit bekannt, in allen tropischen und subtropischen Meeren, nördlich bis zum Mittelmeer, bei Wassertemperaturen zwischen 10 und 32 °C weit verbreitet. Regelmäßig wird Trichoplax adhaerens auch in Meerwasser-Aquarien „gefunden“, so etwa im südenglischen Plymouth oder im US-amerikanischen Bundesstaat Florida in Miami.
Eine direkte Nachsuche im Meer ist für die sehr kleinen und unauffälligen Tiere unmöglich. Zum Nachweis existieren zwei Methoden: Entweder werden Hartsubstrate wie Steine im Lebensraum eingesammelt und später im Aquarium näher untersucht, oder es werden künstliche Substrate, meist Objektträger aus Glas, im Lebensraum exponiert, wobei man auf eine spontane Besiedlung durch die Tiere hofft. Die bisherigen Feldfunde stammen aus Flachwasser der Meeresküsten, bis in eine Tiefe von etwa 20 Meter, und Gezeitenzonen tropischer und subtropischer Meere, wo sich die Tiere auf Substraten wie Stämmen oder Wurzeln von Mangroven, Weichtier-Schalen, Bruchstücken von Steinkorallen oder einfach auf Felsbrocken finden. Sie können sowohl Brackwasser-beeinflusste Lebensräume mit einer Salinität bis 2 % wie auch etwas aufgesalzenes, konzentriertes Meerwasser mit einer Salinität bis über 5 % überleben. Ihre Häufigkeit ist in den Sommermonaten höher.[8] Die Häufigkeit des Vorkommens steigt, nach Modellberechnungen, mit steigender Temperatur und steigender Salinität an, wobei eher nährstoffarmes Wasser bevorzugt wird. Die unterschiedlichen genetischen Linien (die vermutlich verschiedenen Arten entsprechen) zeigen dabei unterschiedliche Vorlieben.[9]
Trichoplax adhaerens ernährt sich von kleinen Algen, insbesondere Grünalgen (Chlorophyta) der Gattung Chlorella, Kryptomonaden (Cryptophyta) der Gattungen Cryptomonas oder Rhodomonas, Cyanobakterien (Cyanobacteria) wie Phormidium inundatum, aber auch von abgestorbenen Teilen anderer Organismen. Dazu bilden sich auf der Bauchseite um die Nahrungspartikel eine oder mehrere kleine Taschen, in die von den Drüsenzellen Verdauungsenzyme abgegeben werden; die Tiere entwickeln also vorübergehend einen „Außenmagen“. Die aufgeschlossenen Nährstoffe werden durch Pinocytose („Zell-Trinken“) von den bauchseitigen Geißelzellen aufgenommen.
Auch über das obere Epitheloid (den „Rücken“ des Tieres) können Nahrungspartikel, auch ganze Einzeller, aufgenommen werden. Diese Ernährungsweise dürfte im Tierreich einmalig sein: In einer Schleimschicht gesammelte Nahrungspartikel werden von Faserzellen durch Interzellular-Spalten (Zellzwischenräume) des Epitheloids gezogen und per Phagocytose („Zell-Essen“) verdaut. Dieses „Einsammeln“ von Nahrungspartikeln durch ein intaktes Abschlussgewebe hindurch ist nur möglich, weil Placozoa verschiedene „abdichtende“ Elemente (eine Basallamina unter dem Epitheloid und bestimmte Zell-Zell-Verbindungen) fehlen.
Mit diesem scheinbar einfachsten Tier leben zwei verschiedene sehr wählerische Bakterien in intrazellulärer Symbiose:
Auf fester Oberfläche können sich Placozoa auf zwei verschiedene Arten bewegen: Einmal ermöglicht es ihnen ihre begeißelte Kriechsohle, langsam über den Untergrund hinweg zu gleiten, zum anderen können sie wie eine Amöbe ihre Lage durch Wandlung der Körperform verändern. Die Bewegungen sind dabei nicht zentral koordiniert, da keinerlei Muskel- oder Nervengewebe existiert.
Zwischen Körperform und Fortbewegungsgeschwindigkeit konnte eine enge Verbindung nachgewiesen werden, die zusätzlich abhängig vom Nahrungsangebot ist[11]
Die beiden Ausbreitungsweisen lassen sich, da der Übergang nicht flüssig, sondern abrupt vonstattengeht, sehr gut gegeneinander abgrenzen. Vereinfachend lässt sich Trichoplax adhaerens als nicht-lineares dynamisches System fern dem thermodynamischen Gleichgewicht modellieren.
Eine qualitative Erklärung für das Verhalten des Tiers ergibt sich wie folgt:
Die konkrete Richtung, in die sich Trichoplax jeweils bewegt, ist zufallsbestimmt: Misst man, wie schnell sich ein einzelnes Tier von einem (willkürlich bestimmten) Ausgangspunkt wegbewegt, so findet man eine lineare Beziehung zwischen der vergangenen Zeit und der mittleren quadratischen Entfernung von Start- und aktuellem Aufenthaltsort. Ein solcher Zusammenhang charakterisiert auch die zufallsbestimmte Brownsche Bewegung, die somit als Modell für die Fortbewegung der Placozoa dienen kann.
Kleine Tiere sind auch in der Lage, aktiv mit Hilfe ihrer Geißeln zu schwimmen. Sobald sie ein mögliches Substrat berühren, kommt es zu einer Dorsoventralreaktion: Die Geißeln der Rückseite schlagen weiter, während die Geißeln der bauchseitigen Zellen ihren Schlagrhythmus einstellen. Gleichzeitig versucht die Bauchseite, Kontakt mit dem Untergrund herzustellen; kleine Zellaus- und -einstülpungen, die Mikrovilli, die sich an der Oberfläche der säuligen Zellen der Bauchseite befinden, tragen durch ihre adhäsive (klebende) Wirkung zur Anheftung an das Substrat bei.
Eine bemerkenswerte Eigenschaft der Placozoa ist, dass sie sich aus kleinsten Zellbeständen regenerieren können. Selbst wenn im Experiment große Teile des Organismus entfernt werden, entwickelt sich aus dem Rest wieder ein vollständiges Tier. Es ist darüber hinaus möglich, Trichoplax adhaerens durch ein Passiersieb zu streichen, so dass die Einzelzellen zwar nicht zerstört, aber weitgehend voneinander getrennt werden. Im Reagenzglas finden sie sich dann wieder zu kompletten Organismen zusammen. Wird diese Prozedur mit mehreren vorher angefärbten Tieren gleichzeitig vollzogen, geschieht das Gleiche. In diesem Fall kann es sogar vorkommen, dass Zellen, die vorher zu einem bestimmten Tier gehörten, plötzlich als Teil eines anderen wieder auftauchen.
Normalerweise pflanzen sich die Placozoa ungeschlechtlich fort. Das Tier schnürt sich dazu in der Mitte durch, so dass zwei etwa gleich große Tochtertiere entstehen, die nach der Fission allerdings noch für eine Weile lose verbunden bleiben. Seltener findet man Knospungsvorgänge: Dabei trennen sich von der Rückenseite kleine, im Wasser schwebende Zellkugeln („Schwärmer“) ab, die alle bekannten Zelltypen in sich vereinigen und in der Folge zu einem eigenen Individuum heranwachsen.
Die geschlechtliche Fortpflanzung wird möglicherweise durch eine zu große Populationsdichte, in einigen Fällen durch hohe Wassertemperaturen, ausgelöst, sie ist bisher rätselhaft und im Detail kaum verstanden. Im Labor bilden dabei ausschließlich degenerierende, durch Wasseraufnahme anschwellende Individuen binnen vier bis sechs Wochen Eizellen aus. Diese werden vermutlich aus Zellen des unteren Epithels gebildet und wachsen ins Innere des Organismus aus. Dabei schwellen sie durch Aufnahme von Nährstoffen und (extern gebildetem) Dotter durch Phagocytose aus dem sich auflösenden mütterlichen Organismus bis zu einer Größe von 70–120 µm an. Die meisten Individuen bilden nur eine einzige Eizelle, selten kommen bis zu drei vor. Gleichzeitig gebildete, sogenannte F-Zellen stellen möglicherweise die männlichen Geschlechtszellen (Spermien) dar[12], doch wurde weder die Funktion bestätigt noch der Befruchtungsvorgang bisher beobachtet; allerdings werden einige für Spermien typische Proteine in den Zellen exprimiert. Nach der (angenommenen) Befruchtung bildet die Eizelle die sogenannte Befruchtungsmembran, eine schützende Hülle. Die Embryonalentwicklung beginnt mit einer vollständigen, äqualen Furchung. Die jungen Embryonen wachsen im mütterlichen Organismus heran, sie werden durch dessen Degeneration und Auflösung freigesetzt.[13]
Trotz erheblicher Anstrengungen ist es bisher nie gelungen, Embryonen jenseits des 128-Zell-Stadiums am Leben zu erhalten. Die weitere Entwicklung ist daher unbekannt. Vermutlich fehlt in der Laborumgebung ein bisher unverstandener kritischer Faktor des natürlichen Lebensraums.
Wegen der Möglichkeit, sich unbegrenzt durch ungeschlechtliche Fortpflanzung zu klonen, ist die Lebensspanne der Placozoa potentiell unendlich; in der Praxis werden einige Entwicklungslinien, die auf ein einziges Tier zurückgehen, seit längerem – in einem Fall seit 1969 – ohne Auftreten sexueller Prozesse in Kultur gehalten.[14]
Die stammesgeschichtlichen Verwandtschaftsverhältnisse der Placozoa sind umstritten. Fossile Nachweise existieren nicht und sind auch nicht zu erwarten, so dass die Stellung von Trichoplax adhaerens allein aufgrund des Vergleichs rezenter Arten erschlossen werden muss. Die Placozoa sind neben den Schwämmen (Porifera), den Nesseltieren (Cnidaria), den Rippenquallen (Ctenophora) und den Zweiseitentieren (Bilateria) eine der fünf grundlegenden Entwicklungslinien der vielzelligen Tiere (als Taxon Metazoa genannt). Das Verhältnis dieser fünf Gruppen zueinander ist wissenschaftlich umstritten. Zum Verhältnis von 5 Taxa bestehen theoretisch 105 Möglichkeiten der Abstammungsverhältnisse, zahlreiche davon sind als Hypothese schon vorgeschlagen worden. Die bisher erzielten Ergebnisse, von denen einige als zunächst gut abgesichert erschienen, erwiesen sich als stark von der verwendeten Methode und der Anzahl und Zusammensetzung der jeweils einbezogenen Taxa abhängig und gelten damit alle als unsicher.[15][16]
Die Zuordnung zu den Gewebelosen Tieren („Parazoa“) beruht nicht auf den angenommenen Verwandtschaftsbeziehungen der Placozoa, sondern klassifiziert die Tiere stattdessen nach ihrem Organisationsgrad: Ebenso wie den Schwämmen (Porifera), mit denen sie in diesem Taxon vereinigt sind, fehlen ihnen Gewebe oder Organe; das Epitheloid wird in diesem Zusammenhang nicht als vollwertiges Gewebe angesehen. Allerdings sind dies „primitive Merkmale“, so genannte Symplesiomorphien, die auf die gemeinsame Stammart aller Tiere zurückgehen und daher nach Ansicht der phylogenetischen Systematik keine evolutionäre Verwandtschaft begründen können. Andere Biologen sehen darin Rückbildungen, die Tiere wären also sekundär vereinfachte Nachkommen komplizierter organisierter Vorfahren.
Aufgrund ihres einfachen Aufbaus werden die Placozoa manchmal als Modellorganismen für die Entwicklung von Einzellern zu Vielzellern angesehen. Sie gelten dann als Schwestergruppe aller anderen Vielzeller:
Vielzellige Tiere (Metazoa) N.N.Schwämme (Porifera)
Gewebetiere (Eumetazoa)
Placozoa
Ein wichtiges Argument für eine solche basale Stellung der Placozoa ist, neben ihrer morphologisch einfachen Organisation, der Aufbau der mitochondrialen DNA, des eigenständigen Erbguts der Mitochondrien. Diese ist, wie bei vielen Einzellern, äußerst komplex aufgebaut, weitaus komplizierter als diejenige aller anderen Vielzeller, unter Einschluss der Schwämme.[17] Phylogenomische Studien, bei denen die Verwandtschaftsverhältnisse anhand des Vergleichs homologer DNA-Sequenzen der mtDNA analysiert worden sind, erwiesen sich aber als nicht sehr erhellend.[18] Vermutlich ist die DNA der Mitochondrien aufgrund zu vieler Mutationen nicht mehr phylogenetisch informativ genug. Genetische Studien, die ein Schwestergruppen-Verhältnis der Placozoa zu den anderen Metazoa aufzeigten gelten daher als sehr unsicher und kaum verlässlich.[19]
Das wichtigste, auf rein morphologischen Eigenschaften beruhende Konzept sieht die Placozoa als nächste Verwandte der Gewebetiere (Eumetazoa). Das gemeinsame, als Epitheliozoa bezeichnete Taxon wird seinerseits als Schwestergruppe der Schwämme (Porifera) aufgefasst:[20]
Vielzellige Tiere (Metazoa) EpitheliozoaPlacozoa
Nesseltiere (Cnidaria)
Schwämme (Porifera)
Für ein solches Verhältnis sprechen in erster Linie spezielle Zell-Zell-Verbindungen, die Gürteldesmosomen, die nicht nur bei den Placozoa, sondern bei allen Tieren außerhalb der Schwämme vorkommen und dafür sorgen, dass sich Zellen zu einer lückenlosen Schicht wie dem Epitheloid der Placozoa zusammenschließen können. Das dorsale Epitheloid wird der Hypothese gemäß als homolog zum Ektoderm, das ventrale zum Entoderm der anderen Metazoa aufgefasst. Auch die auf der Bauchseite auftretenden Drüsenzellen teilt Trichoplax adhaerens mit den meisten Gewebetieren. Beide Eigenschaften können als Apomorphien, also als evolutionär abgeleitete Merkmale, gelten und begründen so ein gemeinsames Taxon aller betroffenen Tiere.
Eine dritte, in erster Linie molekulargenetisch begründete Hypothese sieht die Placozoa als stark vereinfachte Gewebetiere. Demnach stammt Trichoplax adhaerens von wesentlich komplexer aufgebauten Tieren ab, die bereits über Muskeln und Nervengewebe verfügten. Beide Gewebetypen wie auch die Basallamina des Epithels sind demzufolge erst durch radikale sekundäre Vereinfachung verloren gegangen.
Verschiedene Studien kommen dabei derzeit zu unterschiedlichen Ergebnissen hinsichtlich der genauen Schwestergruppe: Mal gelten die Placozoa als nächste Verwandte der Nesseltiere (Cnidaria), mal als Schwestergruppe der Rippenquallen und gelegentlich werden sie sogar direkt neben die zweiseitig-symmetrischen Bilateria gestellt:
Vielzellige Tiere (Metazoa) Gewebetiere (Eumetazoa) N.N.Nesseltiere (Cnidaria)
Placozoa
Bilateria
Rippenquallen (Ctenophora)
Schwämme (Porifera)
Lediglich eine ebenfalls vorgeschlagene Einordnung in die Nesseltiere lässt sich derzeit mit hoher Wahrscheinlichkeit ausschließen.
Kritisch wird gegen das vorgeschlagene Szenario eingewandt, dass es morphologische Merkmale der Tiere vollkommen unberücksichtigt lässt. Eine so extreme Vereinfachung, wie sie dem Modell gemäß für die Placozoa postuliert werden müsste, ist zudem nur von parasitisch lebenden Organismen bekannt, ließe sich aber für eine frei lebende Tierart wie Trichoplax adhaerens nur schwer funktional erklären.
Derzeit wird nur eine einzige Art, Trichoplax adhaerens, als Bestandteil der Placozoa anerkannt. Allerdings wurde 1893 durch den Italiener Francesco Saverio Monticelli eine weitere Art mit dem Namen Treptoplax reptans beschrieben, die er in den Gewässern um Neapel fand. Sie gilt jedoch seit 1896 als verschollen; ihre Existenz wird heute von den meisten Zoologen angezweifelt.[1]
Weil zwischen Vertretern von Trichoplax adhaerens oft große genetische Unterschiede bestehen, die bei anderen Organismengruppen zu einer Einteilung in unterschiedliche Gattungen führen würden, erscheint es sehr wahrscheinlich, dass die nach morphologischen Kriterien einzige Art tatsächlich einer Gruppe kryptischer, also äußerlich nicht unterscheidbarer Arten entspricht. Tatsächlich wurden, nach elektronenmikroskopischen Untersuchungen, auch erste morphologische Unterschiede zwischen verschiedenen im Labor kultivierten Klonen angegeben. Bisher sind etwa 200 unterschiedliche genetische Linien, teilweise mit weiter geographischer Verbreitung, nachgewiesen worden, die möglicherweise etwa 20–30 Arten entsprechen könnten. Das Problem der Artabgrenzung bei der merkmalsarmen Gruppe ist aber derzeit noch ungelöst. Auch von der Zuweisung höherer Ränge der rangbasierten (Linné’schen) Taxonomie, wie Familien oder Ordnungen, wird derzeit in der Regel als unzweckmäßig abgesehen.[8]
Trichoplax adhaerens ist die einzige allgemein anerkannte Art der Placozoa. Der Organismus wurde 1883 von dem deutschen Zoologen Franz Eilhard Schulze in einem Meerwasser-Aquarium des Zoologischen Instituts in Graz entdeckt. Der von ihm verliehene Gattungsname des Tiers leitet sich aus dem altgriechischen θρίξ thrix „Haar“ und πλάξ plax „Platte“ ab, das lateinische Artepithet adhaerens bedeutet „haftend“, so dass sich der Artenname etwa mit „anhaftende haarige Platte“ übersetzen lässt.
Die Placozoa oder Plattentiere sind die strukturell einfachsten aller vielzelligen Tiere (Metazoa) und bilden einen eigenen Tierstamm. Der wissenschaftliche Name bedeutet wörtlich übersetzt „flache Tiere“, im Deutschen wird gelegentlich der Name Scheibentiere verwendet. Anhand genetischer Daten gilt es als sicher, dass die Placozoa zahlreiche Arten umfassen. Da sie sich aber morphologisch nicht unterscheiden lassen, wurde bisher nur Trichoplax adhaerens als einzige Art beschrieben.
Lange Zeit hielt man die bisher nicht in ihrem natürlichen Lebensraum beobachteten Tiere für Jugendstadien von Nesseltieren (Cnidaria): Länger als ein halbes Jahrhundert wurden sie als Planula-Larven der Hydrozoen-Art Eleutheria krohni gedeutet. Erst Arbeiten aus den 1970er Jahren, unter anderem durch den Tübinger Protozoologen Karl G. Grell, klärten die frühen Phasen der Embryonalentwicklung der Tiere auf und trugen maßgeblich zur Errichtung eines eigenen Tierstamms für sie bei.
Trichoplax adhaerens is one of the four named species in the phylum Placozoa. The others are Hoilungia hongkongensis, Polyplacotoma mediterranea and Cladtertia collaboinventa. Placozoa is a basal group of multicellular animals, possible relatives of Cnidaria.[2] Trichoplax are very flat organisms around a millimetre in diameter, lacking any organs or internal structures. They have two cellular layers: the top epitheloid layer is made of ciliated "cover cells" flattened toward the outside of the organism, and the bottom layer is made up of cylinder cells that possess cilia used in locomotion, and gland cells that lack cilia.[3] Between these layers is the fibre syncytium, a liquid-filled cavity strutted open by star-like fibres.
Trichoplax feed by absorbing food particles—mainly microbes—with their underside. They generally reproduce asexually, by dividing or budding, but can also reproduce sexually. Though Trichoplax has a small genome in comparison to other animals, nearly 87% of its 11,514 predicted protein-coding genes are identifiably similar to known genes in other animals.
Trichoplax was discovered in 1883 by the German zoologist Franz Eilhard Schulze, in a seawater aquarium at the Zoological Institute in Graz, Austria. The generic name is derived from the classical Greek θρίξ (thrix), "hair", and πλάξ (plax), "plate". The specific epithet adhaerens is Latin meaning "adherent", reflecting its propensity to stick to the glass slides and pipettes used in its examination.[4]
Although from the very beginning most researchers who studied Trichoplax in any detail realized that it had no close relationship to other animal phyla, the zoologist Thilo Krumbach published a hypothesis that Trichoplax is a form of the planula larva of the anemone-like hydrozoan Eleutheria krohni in 1907. Although this was refuted in print by Schulze and others, Krumbach's analysis became the standard textbook explanation, and nothing was printed in zoological journals about Trichoplax until the 1960s. In the 1960s and 1970s a new interest among researchers led to acceptance of Placozoa as a new animal phylum. Among the new discoveries was study of the early phases of the animals' embryonic development and evidence that the animals that people had been studying are adults, not larvae. This newfound interest also included study of the organism in nature (as opposed to aquariums).[5]

Trichoplax generally has a thinly flattened, plate-like body in cross-section around half a millimetre, occasionally up to two or three millimetres. The body is usually only about 25 µm thick. These colorlessly gray organisms are so thin they are transparent when illuminated from behind, and in most cases are barely visible to the naked eye. Like the single-celled amoebae, which they superficially resemble, they continually change their external shape. In addition, spherical phases occasionally form. These may facilitate movement to new habitats.
Trichoplax lacks tissues and organs; there is also no manifest body symmetry, so it is not possible to distinguish anterior from posterior or left from right. It is made up of a few thousand cells of six types in three distinct layers: dorsal epithelia cells and ventral epithelia cells, each with a single cilium ("monociliate"), ventral gland cells, syncytial fiber cells, lipophils, and crystal cells (each containing a birefringent crystal, arrayed around the rim). Lacking sensory and muscle cells, it moves using cilia on its external surface.[6]
There are no neurons present, but in the absence of a nervous system the animal uses short chains of amino acids known as peptides for cell communication, in a manner resembling the way animals with neurons use neuropeptides for the same purpose. Individual cells contain and secrete a variety of small peptides, made up of between four and 20 amino acids, which are detected by neighbouring cells. Each peptide can be used individually to send a signal to other cells, but also sequentially or together in different combinations, creating a huge number a different types of signals. This allows for a relatively complex behavioural repertoire, including behaviours such as "crinkling", turning, flattening, and internal "churning".[7]
Both structurally and functionally, it is possible to distinguish a back or dorsal side from a belly or ventral side in Trichoplax adhaerens. Both consist of a single layer of cells coated on the outside with slime and are reminiscent of epithelial tissue, primarily due to the junctions—belt desmosomes—between the cells. In contrast to true epithelium, however, the cell layers of the Placozoa possess no basal lamina, which refers to a thin layer of extracellular material underlying epithelium that stiffens it and separates it from the body's interior. The absence of this structure, which is otherwise to be found in all animals except the sponges, can be explained in terms of function: a rigid separating layer would make the amoeboid changes in the shape of Trichoplax adhaerens impossible. Instead of an epithelium, therefore, we speak of an epitheloid in the Placozoa.
A mature individual consists of up to a thousand cells that can be divided into four different cell types. The monociliated cells of the dorsal epitheloid are flattened and contain lipid bodies. The cells on the ventral side likewise possess a single cilium, while their elongated columnar shape, with a small cross section at the surface, packs them very closely together, causing the cilia to be very closely spaced on the ventral side and to form a ciliated "crawling sole". Interspersed among these ventral epithlioid cells are unciliated gland cells thought to be capable of synthesizing digestive enzymes.
Between the two layers of cells is a liquid-filled interior space, which, except for the immediate zones of contact with the ventral and dorsal sides, is pervaded by a star-shaped fibre syncytium: a fibrous network that consists essentially of a single cell but contains numerous nuclei that, while separated by internal crosswalls (septa), do not have true cell membranes between them. Similar structures are also found in the sponges (Porifera) and many fungi.
On both sides of the septa are liquid-filled capsules that cause the septa to resemble synapses, i.e. nerve-cell junctions that occur in fully expressed form only in animals with tissues (Eumetazoa). Striking accumulations of calcium ions, which may have a function related to the propagation of stimuli, likewise suggest a possible role as protosynapses. This view is supported by the fact that fluorescent antibodies against cnidarian neurotransmitters, i.e. precisely those signal carriers that are transferred in synapses, bind in high concentrations in certain cells of Trichoplax adhaerens, and thus indicate the existence of comparable substances in the Placozoa. The fibre syncytium also contains molecules of actin and probably also of myosin, which occur in the muscle cells of eumetazoans. In the placozoans, they ensure that the individual fibres can relax or contract and thus help determine the animals' shape.
In this way, the fibre syncytium assumes the functions of nerve and muscle tissues. Moreover, at least a portion of digestion occurs here. On the other hand, no gelatinous extracellular matrix exists of the kind observed, in mesoglea, in cnidarians and ctenophores.
Pluripotent cells, which can differentiate into other cell types, have not yet been demonstrated unambiguously in T. adhaerens, in contrast to the case of the Eumetazoa. The conventional view is that dorsal and ventral epithelioid cells arise only from other cells of the same type.
The Trichoplax genome contains about 98 million base pairs and 11,514 predicted protein-coding genes.[8]
All nuclei of placozoan cells contain six pairs of chromosomes that are only about two to three micrometres in size. Three pairs are metacentric, meaning that the centromere, the attachment point for the spindle fibers in cell division, is located at the center, or acrocentric, with the centromere at an extreme end of each chromosome. The cells of the fiber syncytium can be tetraploid, i.e. contain a quadruple complement of chromosomes.
A single complement of chromosomes in Trichoplax adhaerens contains a total of fewer than fifty million base pairs and thus forms the smallest animal genome; the number of base pairs in the intestinal bacterium Escherichia coli is smaller by a factor of only ten.
The genetic complement of Trichoplax adhaerens has not yet been very well researched; it has, however, already been possible to identify several genes, such as Brachyury and TBX2/TBX3, which are homologous to corresponding base-pair sequences in eumetazoans. Of particular significance is Trox-2, a placozoan gene known under the name Cnox-2 in cnidarians and as Gsx in the bilaterally symmetrical Bilateria. As a homeobox or Hox gene it plays a role in organization and differentiation along the axis of symmetry in the embryonic development of eumetazoans; in cnidarians, it appears to determine the position of mouth-facing (oral) and opposite-facing (aboral) sides of the organism. Since placozoans possess no axes of symmetry, exactly where the gene is transcribed in the body of Trichoplax is of special interest. Antibody studies have been able to show that the gene's product occurs only in the transition zones of the dorsal and ventral sides, perhaps in a fifth cell type that has not yet been characterized. It is not yet clear whether these cells, contrary to traditional views, are stem cells, which play a role in cell differentiation. In any case, Trox-2 can be considered a possible candidate for a proto-Hox gene, from which the other genes in this important family could have arisen through gene duplication and variation.
Initially, molecular-biology methods were applied unsuccessfully to test the various theories regarding Placozoa's position in the Metazoa system. No clarification was achieved with standard markers such as 18S rDNA/RNA: the marker sequence was apparently "garbled", i.e. rendered uninformative as the result of many mutations. Nevertheless, this negative result supported the suspicion that Trichoplax might represent an extremely primitive lineage of metazoans, since a very long period of time had to be assumed for the accumulation of so many mutations.
Of the 11,514 genes identified in the six chromosomes of Trichoplax, 87% are identifiably similar to genes in cnidarians and bilaterians. In those Trichoplax genes for which equivalent genes can be identified in the human genome, over 80% of the introns (the regions within genes that are removed from RNA molecules before their sequences are translated in protein synthesis) are found in the same location as in the corresponding human genes. The arrangement of genes in groups on chromosomes is also conserved between the Trichoplax and human genomes. This contrasts to other model systems such as fruit flies and soil nematodes that have experienced a paring down of non-coding regions and a loss of the ancestral genome organizations.[9]
The phylogenetic relationship between Trichoplax and other animals has been debated for some time. A variety of hypotheses have been advanced based on the few morphological characteristics of this simple organism that could be identified. More recently, a comparison of the Trichoplax mitochondrial genome suggested that Trichoplax is a basal metazoan—less closely related to all other animals including sponges than they are to each other.[10] This implies that the Placozoa would have arisen relatively soon after the evolutionary transition from unicellular to multicellular forms. But an even more recent analysis of the much larger Trichoplax nuclear genome instead supports the hypothesis that Trichoplax is a basal eumetazoan, that is, more closely related to Cnidaria and other animals than any of those animals are to sponges.[8] This is consistent with the presence in Trichoplax of cell layers reminiscent of epithelial tissue (see above).
Trichoplax was first discovered on the walls of a marine aquarium, and is rarely observed in its natural habitat.[11] Trichoplax has been collected, among other places, in the Red Sea, the Mediterranean, and the Caribbean, off Hawaii, Guam, Samoa, Japan, Vietnam, Brazil, and Papua New Guinea, and on the Great Barrier Reef off the east coast of Australia.[12]
Field specimens tend to be found in the coastal tidal zones of tropical and subtropical seas, on such substrates as the trunks and roots of mangroves, shells of molluscs, fragments of stony corals or simply on pieces of rock. One study was able to detect seasonal population fluctuations, the causes of which have not yet been deduced.
Trichoplax adhaerens feeds on small algae, particularly on green algae (Chlorophyta) of the genus Chlorella, cryptomonads (Cryptophyta) of the genera Cryptomonas and Rhodomonas, and blue-green bacteria (Cyanobacteria) such as Phormidium inundatum, but also on detritus from other organisms. In feeding, one or several small pockets form around particles of nutrients on the ventral side, into which digestive enzymes are released by the gland cells; the organisms thus develop a temporary "external stomach", so to speak. The enclosed nutrients are then taken up by pinocytosis ("cell-drinking") by the ciliated cells located on the ventral surface.
Entire single-celled organisms can also be ingested through the upper epitheloid (that is, the "dorsal surface" of the animal). This mode of feeding could be unique in the animal kingdom: the particles, collected in a slime layer, are drawn through the intercellular gaps (cellular interstices) of the epitheloid by the fibre cells and then digested by phagocytosis ("cell-eating"). Such "collecting" of nutrient particles through an intact tegument is only possible because some "insulating" elements (specifically, a basal lamina under the epitheloid and certain types of cell-cell junctions) are not present in the Placozoa.
Not all bacteria in the interior of Placozoa are digested as food: in the endoplasmic reticulum, an organelle of the fibre syncytium, bacteria are frequently found that appear to live in symbiosis with Trichoplax adhaerens.[13]
Placozoa can move in two different ways on solid surfaces: first, their ciliated crawling sole lets them glide slowly across the substrate; second, they can change location by modifying their body shape, as an amoeba does. These movements are not centrally coordinated, since no muscle or nerve tissues exist. It can happen that an individual moves simultaneously in two different directions and consequently divides into two parts.[14]
It has been possible to demonstrate a close connection between body shape and the speed of locomotion, which is also a function of available food:
Since the transition is not smooth but happens abruptly, the two modes of extension can be very clearly separated from one another. The following is a qualitative explanation of the animal's behavior:
The actual direction in which Trichoplax moves each time is random: if we measure how fast an individual animal moves away from an arbitrary starting point, we find a linear relationship between elapsed time and mean square distance between starting point and present location. Such a relationship is also characteristic of random Brownian motion of molecules, which thus can serve as a model for locomotion in the Placozoa.
Small animals are also capable of swimming actively with the aid of their cilia. As soon as they come into contact with a possible substrate, a dorsoventral response occurs: the dorsal cilia continue to beat, whereas the cilia of ventral cells stop their rhythmic beating. At the same time, the ventral surface tries to make contact with the substrate; small protrusions and invaginations, the microvilli found on the surface of the columnar cells, help in attaching to the substrate via their adhesive action.
Using Trichoplax adhaerens as a model, were described 0.02–0.002 Hz oscillations in locomotory and feeding patterns as evidence of complex multicellular integration; and showed their dependence on the endogenous secretion of signal molecules. Evolutionary conserved low-molecular-weight transmitters (glutamate, aspartate, glycine, GABA, and ATP) acted as coordinators of distinct locomotory and feeding patterns. Specifically, L-glutamate induced and partially mimicked endogenous feeding cycles, whereas glycine and GABA suppressed feeding. ATP-modified feeding is complex, first causing feeding-like cycles and then suppressing feeding. Trichoplax locomotion was modulated by glycine, GABA, and, surprisingly, by animals’ own mucus trails. Mucus triples locomotory speed compared to clean substrates. Glycine and GABA increased the frequency of turns. [15]
A notable characteristic of the Placozoa is that they can regenerate themselves from extremely small groups of cells. Even when large portions of the organism are removed in the laboratory, a complete animal develops again from the remainder. It is also possible to rub Trichoplax adhaerens through a strainer in such a manner that individual cells are not destroyed but are separated from one another to a large extent. In the test tube they then find their way back together again to form complete organisms. If this procedure is performed on several previously strained individuals simultaneously, the same thing occurs. In this case, however, cells that previously belonged to a particular individual can suddenly show up as part of another.
The Placozoa normally propagate asexually, dividing down the middle to produce two (or sometimes, three) roughly equal-sized daughters. These remain loosely connected for a while after fission. More rarely, budding processes are observed: spherules of cells separate from the dorsal surface; each of these combines all known cell types and subsequently grows into an individual on its own.
Sexual reproduction is thought to be triggered by excessive population density. As a result, the animals absorb liquid, begin to swell, and separate from the substrate so that they float freely in the water. In the protected interior space, the ventral cells form an ovum surrounded by a special envelope, the fertilisation membrane; the ovum is supplied with nutrients by the surrounding syncytium, allowing energy-rich yolk to accumulate in its interior. Once maturation of the ovum is complete, the rest of the animal degenerates, liberating the ovum itself. Small, unciliated cells that form at the same time are interpreted to be spermatozoa. It has not yet been possible to observe fertilisation itself; the existence of the fertilisation membrane is currently taken to be evidence, however, that it has taken place.
Putative eggs have been observed, but they degrade, typically at the 32–64 cell stage. Neither embryonic development nor sperm have been observed. Despite lack of observation of sexual reproduction in the lab, the genetic structure of the populations in the wild is compatible with the sexual reproduction mode, at least for species of the analysed genotype H5.[16]
Usually even before its liberation, the ovum initiates cleavage processes in which it becomes completely pinched through at the middle. A ball of cells characteristic of animals, the blastula, is ultimately produced in this manner, with a maximum of 256 cells. Development beyond this 256-cell stage has not yet been observed.[17]
Trichoplax lack a homologue of the Boule protein that appears to be ubiquitous and conserved in males of all species of other animals tested.[18] If its absence implies the species has no males, then perhaps its "sexual" reproduction may be a case of the above-described process of regeneration, combining cells from two separate organisms into one.
Due to the possibility of its cloning itself by asexual propagation without limit, the life span of Placozoa is infinite; in the laboratory, several lines descended from a single organism have been maintained in culture for an average of 20 years without the occurrence of sexual processes.
Long ignored as an exotic, marginal phenomenon, Trichoplax adhaerens is today viewed as a potential biological model organism. In particular, research is needed to determine how a group of cells that cannot be considered full-fledged epithelial tissue organizes itself, how locomotion and coordination occur in the absence of true muscle and nerve tissue, and how the absence of a concrete body axis affects the animal's biology. At the genetic level, the way in which Trichoplax adhaerens protects against damage to its genome needs to be studied, particularly with regard to the existence of special DNA-repair processes. T. adhaerens can tolerate high levels of radiation damage that are lethal to other animals.[19] Tolerance to X-ray exposure was found to depend on expression of genes involved in DNA repair and apoptosis including the gene Mdm2.[19] Complete decoding of the genome should also clarify the placozoans' place in evolution, which continues to be controversial.
Its ability to fight cancer through a combination of aggressive DNA repair and ejection of damaged cells makes it a promising organism for cancer research.[20]
In addition to basic research, this animal could also be suitable for studying wound-healing and regeneration processes; as yet unidentified metabolic products should be researched. Finally, Trichoplax adhaerens is also being considered as an animal model for testing compounds and antibacterial drugs.[21]
Francesco Saverio Monticelli described another species in 1893, which he found in the waters around Naples, naming it Treptoplax reptans. However, it has not been observed since 1896, and most zoologists today doubt its existence.
Significant genetic differences have been observed between collected specimens matching the morphological description of T. adhaerens, suggesting that it may be a cryptic species complex. At least eight distinct genotypes (marked from H1 to H8) have been observed.[22]
Because great genetic differences often occur between representatives of Trichoplax adhaerens, differences that in other taxa would result in their being spread among different genera, it is currently unclear whether the single species, based on morphological criteria, does not actually correspond to a group of cryptospecies, i.e. species that are not outwardly distinguishable from one another. Distribution of the genetic variants is not a function of geography: some variants are found in multiple regions (e.g. Pacific, Caribbean and Red Sea). At the same time, very different genetic variants can be isolated from the same habitat.
{{cite journal}}: CS1 maint: multiple names: authors list (link) {{cite web}}: CS1 maint: url-status (link) {{cite journal}}: CS1 maint: multiple names: authors list (link) Trichoplax adhaerens is one of the four named species in the phylum Placozoa. The others are Hoilungia hongkongensis, Polyplacotoma mediterranea and Cladtertia collaboinventa. Placozoa is a basal group of multicellular animals, possible relatives of Cnidaria. Trichoplax are very flat organisms around a millimetre in diameter, lacking any organs or internal structures. They have two cellular layers: the top epitheloid layer is made of ciliated "cover cells" flattened toward the outside of the organism, and the bottom layer is made up of cylinder cells that possess cilia used in locomotion, and gland cells that lack cilia. Between these layers is the fibre syncytium, a liquid-filled cavity strutted open by star-like fibres.
Trichoplax feed by absorbing food particles—mainly microbes—with their underside. They generally reproduce asexually, by dividing or budding, but can also reproduce sexually. Though Trichoplax has a small genome in comparison to other animals, nearly 87% of its 11,514 predicted protein-coding genes are identifiably similar to known genes in other animals.
Trichoplax adhaerens est une espèce de placozoaires.
Le génome de Trichoplax adhaerens comporte environ 98 millions de paires de bases et 11 500 gènes, dont près de 87 % ont leurs correspondants chez d'autres animaux[1].
L'organisme de Trichoplax adhaerens est constitué de deux couches reliées par des cellules en forme de fibre. On dénombre au total six types cellulaires dont notamment, dans la couche ventrale, des cellules glandulaires (secrétant des molécules servant à la communication), des cellules lipophiles (contenant des granules huileux) et des cellules épithéliales ciliées[1].
Découvert en 1883, Trichoplax adhaerens est considéré jusqu'en 2018 comme l'unique espèce de l'embranchement des Placozoaires. En 2018 on découvre une autre espèce (Hoilungia hongkongensis[2]) placée dans la même famille (Trichoplacidae), puis en 2019 une troisième (Polyplacotoma mediterranea) placée dans une seconde famille (non encore nommée)[3].
Trichoplax adhaerens est une espèce de placozoaires.
O Trichoplax adhaerens Schultze é unha especie animal que pertence ao filo dos placozoos, que ten forma de globo, cunha cavidade corporal enchida de fluído a presión.
É un animal mariño diminuto, que repta sobre o substrato cos seus cilios, e está composto de capas epiteliais ventrais e dorsais, contendo células mesenquimáticas frouxas. A reprodución é sexual, pero pode ser tamén asexual, por fisión e por xemación.
É o único representante do filo dos placozoos, xa que outra especie (Treptoplax reptans) foi descrita en 1896 e dende entón xamais foi rexistada, levantando dúbidas sobre a súa real existencia.
A súas relacións evolutivas están sendo investigadas, pero podería estar próximas aos cnidarios e aos ctenóforos. Dellaporta et al. investigaron a secuencia mitocondrial completa do Trichoplax adhaerens e amosaron que os placozoos son a rama que xurdiu antes dentro dos metazoos.[1]
O Trichoplax adhaerens Schultze é unha especie animal que pertence ao filo dos placozoos, que ten forma de globo, cunha cavidade corporal enchida de fluído a presión.
Trichoplax adhaerens is een plakdiertjessoort uit de familie van de Trichoplacidae. De wetenschappelijke naam van de soort is voor het eerst geldig gepubliceerd in 1883 door Schulze.[1]
Bronnen, noten en/of referentiesTrichoplax adhaerens er eit lite, særs primitivt dyr som finst i marine botnsubstrat over heile verda. Arten manglar symmetri og består av nokre få tusen celler. Cellene er ikkje vidare differensiert utover eit skilje mellom over- og undersida, og er vanlegvis mellom 0,5 og 3 millimeter i diameter.
Kroppsplanet til Trichoplax er enkelt; han består av to celleplater med eit holrom mesenkymet i mellom. Cellene i det øvste laget er avflata og bèr eit flimmerhår kvar. I tillegg inneheldt dei små dropar med olje. Det nedste laget har òg eit flimmerhår per celle, men cellene her er meir sylindriske og manglar olje. Dei kan dessutan danna små innsnøringar for å ta opp det ekstracellulære materialet dyra lever av.
Mellom «rygg»- og «buk»-laga har Trichoplax eit mesenkym med amøbe-liknande celler som flyttar seg rundt i ein geleaktig støttestruktur.
Trichoplax brukar flimmerhåra på buken til å krypa over underlaget, på same måte som tøffeldyr og mange flatmakk. På same måte som amøbar kan dei dessutan til ein viss grad flytta på seg ved å endra kroppsform. Små individ kan i tillegg symja.
Dei tek opp mat gjennom fagocytose, det vil seie at einskildcellene lukkar seg rundt partiklar og bryt dei ned individuelt. Føda deira er hovudsakleg organisk detritus, mikroalgar, protozoar og bakteriar. Næringa dei tek opp vert sannsynlegvis transportert rundt i kroppen via mesenkymet.
Trichoplax formeirar seg både aseksuelt og kjønna. Egga utviklar seg i mesenkymlaget, men me veit ikkje korleis dei vert danna. Kromosoma deira er særs små og inneheldt lite informasjon.
Trichoplax er eit særs primitivt dyr, og det har vorte spekulert ein del i kvar ein skal plassera dei systematisk. Grell (1982) meiner dei ulike cellene i det øvste og det nedste laget tilsvarar dei endo- og ektoderme cellelaga me finn hjå dei fleste fleircella dyr unnateke svampar. Dette vil plassera gruppa mellom svampar og nesledyr i stamtreet til dyreriket. Undersøkingar med mitokondrielt DNA (Dellaporta et al, 2006) støttar òg til ein viss grad dette synet.
Det er likevel langt frå semje om spørsmålet.
I 2008 vart genomet til Trichoplax kartlagt for første gong. Mellom resultata av undersøkinga er mellom anna at organismen har det minste genomet av dei fleircella dyra, og at mange grunnleggjande biokjemiske prosessar er dei same i han som i andre dyr. Over 80 prosent av gena til Trichoplax finst òg i det menneskelege genomet.
Trichoplax adhaerens er eit lite, særs primitivt dyr som finst i marine botnsubstrat over heile verda. Arten manglar symmetri og består av nokre få tusen celler. Cellene er ikkje vidare differensiert utover eit skilje mellom over- og undersida, og er vanlegvis mellom 0,5 og 3 millimeter i diameter.
Trichoplax adhaerens este unicul reprezentant viu al încrengăturii Placozoa, măsurând circa 3 mm în diametru. Trăiește în apa mării și seamănă cu o clătită mică, alcătuită dintr-o mulțime de celule. Se târăște pe stânci, digerând orice algă întâlnită în cale. Deși mai complex decât un protozor, trichoplax nu are organe specializate. A fost descoperit în 1883 într-un acvariu din Austria și, inițial, s-a crezut că ar fi vorba despre o larvă.
Trichoplax adhaerens este unicul reprezentant viu al încrengăturii Placozoa, măsurând circa 3 mm în diametru. Trăiește în apa mării și seamănă cu o clătită mică, alcătuită dintr-o mulțime de celule. Se târăște pe stânci, digerând orice algă întâlnită în cale. Deși mai complex decât un protozor, trichoplax nu are organe specializate. A fost descoperit în 1883 într-un acvariu din Austria și, inițial, s-a crezut că ar fi vorba despre o larvă.
Trichoplax adhaerens là đại diện còn tồn tại duy nhất của ngành Placozoa - một nhóm cơ sở của động vật đa bào (Metazoa). Trichoplax là những sinh vật rất phẳng có đường kính 1 mm, không có bất kỳ bộ phận cơ thể hoặc cấu trúc bên trong. Chúng có hai lớp tế bào: lớp biểu mô (epithelioid) trên gồm các "tế bào che phủ" có lông rung phẳng về phía bên ngoài của sinh vật, và lớp dưới được tạo thành từ các tế bào hình trụ có lông rung (tiêm mao) được sử dụng trong vận động, và các tế bào tuyến thiếu lông rung.[2] Giữa các lớp là hợp bào sợi, một khoang chứa chất lỏng mở bởi những sợi hình sao.
Trichoplax ăn bằng cách hấp thụ thức ăn - chủ yếu là vi sinh vật - bằng mặt dưới của chúng. Chúng thường sinh sản vô tính, bằng cách chia đôi hoặc phân chồi, nhưng cũng có thể sinh sản hữu tính. Mặc dù Trichoplax có bộ gen nhỏ so với các động vật khác, gần 87% của 11.514 gen dự đoán là đồng nhất với các loài động vật khác.
Trichoplax được nhà động vật học người Đức Franz Eilhard Schulze phát hiện năm 1883 trong một bể cá biển tại Viện động vật học ở Graz, Áo. Tên chi có nguồn gốc từ tiếng Hy Lạp θρίξ (thrix), "lông", và πλάξ (plax), "tấm". Ý nghĩa tên gọi riêng xuất phát từ tiếng Latin "dính chặt", phản ánh xu hướng bám dính của nó vào các tấm kính mang vật và ống hút được sử dụng trong kiểm tra.[3]
Mặc dù từ rất sớm, hầu hết các nhà nghiên cứu khi nghiên cứu Trichoplax đều nhận ra rằng nó không có mối quan hệ gần với các ngành động vật khác, nhưng nhà động vật học Thilo Krumbach lại công bố một giả thuyết cho rằng Trichoplax là một hình thức của ấu trùng planula của loài thủy tức Eleutheria krohni trông giống như hải quỳ vào năm 1907. Mặc dù điều này đã bị Schulze và những người khác bác bỏ, phân tích của Krumbach đã trở thành lời giải thích chuẩn của sách giáo khoa, và không có gì được in trên các tạp chí động vật về Trichoplax cho đến thập niên 1960. Trong năm 1960 và 1970 một sự quan tâm mới của các nhà nghiên cứu dẫn đến sự chấp nhận Placozoa như một ngành động vật mới. Trong số những khám phá mới là nghiên cứu về giai đoạn đầu của sự phát triển phôi và bằng chứng cho thấy loài động vật mà người ta đã nghiên cứu là sinh vật trưởng thành, không phải ấu trùng. Sự chú ý mới được phát hiện này cũng bao gồm nghiên cứu về sinh vật trong tự nhiên.[4]
Trichoplax thường có một cơ thể dạng tấm mỏng dẹt tiết diện khoảng 0,5 mm, đôi khi lên đến 2 hoặc 3 mm. Cơ thể thường chỉ dày khoảng 25 µm. Những sinh vật màu xám ít màu sắc này vì thế là trong suốt khi được chiếu sáng từ phía sau, và trong nhiều trường hợp hầu như không thể nhìn thấy bằng mắt thường. Giống như amip đơn bào, mà bề ngoài chúng trông giống, chúng liên tục thay đổi hình dạng bên ngoài. Ngoài ra, thỉnh thoảng chúng có dạng hình cầu. Điều này có thể tạo điều kiện cho sự di chuyển tới môi trường sống mới.
Trichoplax thiếu mô và cơ quan; cũng không có biểu hiện đối xứng cơ thể, vì vậy nó không thể phân biệt trước sau hoặc trái phải. Nó được tạo thành từ vài nghìn tế bào thuộc bốn loại thành ba lớp riêng biệt: các tế bào biểu mô lưng và bụng với một lông rung duy nhất, các tế bào tuyến bụng và các tế bào hợp bào sợi. Nó không có các tế bào giác quan hoặc cơ; nó di chuyển bằng cách sử dụng lông rung trên bề mặt bên ngoài của nó.
Trichoplax adhaerens là đại diện còn tồn tại duy nhất của ngành Placozoa - một nhóm cơ sở của động vật đa bào (Metazoa). Trichoplax là những sinh vật rất phẳng có đường kính 1 mm, không có bất kỳ bộ phận cơ thể hoặc cấu trúc bên trong. Chúng có hai lớp tế bào: lớp biểu mô (epithelioid) trên gồm các "tế bào che phủ" có lông rung phẳng về phía bên ngoài của sinh vật, và lớp dưới được tạo thành từ các tế bào hình trụ có lông rung (tiêm mao) được sử dụng trong vận động, và các tế bào tuyến thiếu lông rung. Giữa các lớp là hợp bào sợi, một khoang chứa chất lỏng mở bởi những sợi hình sao.
Trichoplax ăn bằng cách hấp thụ thức ăn - chủ yếu là vi sinh vật - bằng mặt dưới của chúng. Chúng thường sinh sản vô tính, bằng cách chia đôi hoặc phân chồi, nhưng cũng có thể sinh sản hữu tính. Mặc dù Trichoplax có bộ gen nhỏ so với các động vật khác, gần 87% của 11.514 gen dự đoán là đồng nhất với các loài động vật khác.
Уплощённое, лишённое симметрии[13] тело Trichoplax достигает 2—3 мм в диаметре и всего лишь 25 мкм в толщину. В теле Trichoplax выделяют всего 4 типа клеток, организованных в три слоя: верхний и нижний слои клеток, а также разделяющая их соединительная ткань. Наружный слой клеток очень похож на эпителий из-за наличия типичных клеточных контактов между соседними клетками (ленточных десмосом в апикальной области клеток, связанных с пучками актиновых филаментов, а также контактов, похожих на септированные десмосомы), поэтому он больше напоминает эпителий Metazoa, чем пинакодерму[en] губок. Однако его не подстилает базальная пластинка[en], как все настоящие эпителии. Клетки верхней (дорсальной) поверхности плоские, несут по одной ресничке, их ядросодержащие части вдаются вглубь тела. В верхнем слое клеток располагаются так называемые блестящие шары (нем. Glanzkugeln — термин, введённый Ф. Шульце в 1891 году[4]), которые представляют собой сильно преломляющие свет липидные тельца и являются остатками дегенерировавших клеток (вероятно, они происходят из клеток срединного волокнистого синцития, мигрировавших в верхний слой). Реснички располагаются на дне структурно оформленного углубления. От базального тельца каждой реснички отходят микротрубочки и пучки поперечно-исчерченных фибрилл, обеспечивающих опору реснички[14][15].
Нижняя (вентральная) поверхность обращена к субстрату, покрывающие её клетки представлены безжгутиковыми железистыми клетками и клетками, несущими по одной ресничке. Прочное прикрепление к субстрату (отсюда видовой эпитет adhaerens) обеспечивается выростами ресничных клеток, напоминающими микроворсинки. Эти клетки высокие и узкие, поэтому реснички располагаются близко друг к другу, благодаря чему на нижней поверхности тела животного образуется плотный ресничный покров, обеспечивающий локомоцию[14][16].
Пространство между верхним и нижнем слоями клеток заполнено соединительной тканью. Она представлена волокнистым синцитием, образующим сложную трёхмерную сеть; он залегает в жидкой составляющей, близкой по составу к морской воде из-за более слабых межклеточных контактов, чем у других низших многоклеточных. Её нельзя считать настоящим внеклеточным матриксом, характерным для всех остальных многоклеточных животных, потому что в ней отсутствует коллаген-протеогликан-гликопротеиновый комплекс. Многочисленные ядра мезенхимного волокнистого синцития отделяются друг от друга не мембранами, а внутриклеточными перегородками (септами). Аналогичные септы имеются в синцитиях стеклянных губок и грибов. Впрочем, по последним данным синцитиальное строение сети волокнистых клеток не подтверждается чётко[17]. Ядра этих клеток тетраплоидны, в отличие от диплоидных ядер клеток верхнего и нижнего эпителиев. Их митохондрии связаны в единый комплекс, расположенный вблизи ядра; этот комплекс включает также пузырьки неизвестной природы. Кроме того, волокнистые клетки содержат эндосимбиотические бактерии и, в отличие от клеток верхнего и нижнего слоёв, способны к фагоцитозу. Считается, что волокнистый синцитий сократим, он содержит актин, микротрубочки и, вероятно, миозин. Вероятно, волокнистые клетки функционально соответствуют как мышечным, так и нервным клеткам. Кроме волокнистого синцития, в промежуточном слое залегают половые клетки[14][18].
При росте тела трихоплакса митозы происходят во всех трёх слоях клеточных слоях, причём клетки каждого типа, кроме железистых образуются из себе подобных (железистые клетки образуются от ресничных). Мультипотентные клетки[en] у трихоплакса отсутствуют. Эксперименты по регенерации у трихоплакса показали наличие некоторой степени дифференцировки клеток. Так, из изолированной центральной области пластинки и краевого валика шириной около 20 мкм, образованного несколько более мелкими клетками, восстановление цельной пластинки невозможно. При искусственном соединении краевого и центрального фрагментов происходит отторжение лишнего материала, что свидетельствует о существовании определённого баланса между клетками краевой и центральной частей пластинок. Возможно, процесс деления трихоплакса запускается нарушением этого баланса. При несбалансированном росте обоих слоёв клеток проявляются определённые аномалии строения. Так, при недостатке или полном отсутствии дорсального слоя клеток формируются крупные полые шары из вентрального слоя клеток, который проявляет способность к пиноцитозу и изнутри выстилается волокнистыми клетками. Если же отсутствует вентральный эпителий, то формируются сплошные шары из дорсального эпителия, заполненные волокнистым синцитием[19].
В 2014 году был проведён пересмотр клеточного состава Trichoplax. Согласно новейшим представлениям, в теле трихоплакса выделяют 6 типов соматических клеток: в вентральном слое клеток — ресничные эпителиальные клетки, новоидентифицированные липофильные клетки, заполненные липидными гранулами, и железистые клетки; в промежуточном слое — волокнистые клетки; в дорсальном слое клеток — дорсальные эпителиальные клетки, новоидентифицированные кристаллические клетки. Функциональные и другие особенности типов клеток трихоплакса перечислены в таблице ниже[17].
Типы клеток Trichoplax adhaerens Тип Описание[17] Вентральные эпителиальные клетки Мелкие клетки, вытянутые в дорсовентральном направлении, имеют единственную ресничку, поднимающуюся из особого углубления, и множество микроворсинок. Содержат одно крупное (~ 1 мкм) хлопьевидное включение рядом с ядром, плотные гранулы диаметром 200—400 нм вблизи основания, и мелкие пустые везикулы по всей клетке. Вентральные эпителиальные клетки соединены десмосомами в апикальной области, однако характерные для Metazoa плотные контакты и базальная пластинка отсутствуют. Эти клетки можно визуализировать при помощи флуоресцентных антител к мембранно-связанными гуанилаткиназам (MAGUKs), которые часто ассоциированы с клеточными контактами. На долю вентральных эпителиальных клеток приходится 72 % всех клеток в теле трихоплакса. Липофильные клетки Клетки, лишённые ресничек, разбросаны между вентральными эпителиальными клетками. Содержат крупное (~3 мкм) окружённое мембраной сферическое включение с липидным содержимым, расположенное вблизи вентральной поверхности. Под сканирующим электронными микроскопом липофильные клетки легко отличимы от вентральных эпителиальных клеток благодаря крупным размерам, более глубокому расположению и бугристым очертаниям. Липофильные клетки распределены примерно равномерно в вентральном слое клеток, однако полностью отсутствуют в дорсальном слое. На их долю приходится 11 % клеток в теле трихоплакса, что делает их вторым по многочисленности типом клеток этого организма. Железистые клетки Клетки вентрального эпителия, как предполагается, выполняют секреторную функцию. Содержат многочисленные гранулы размером 200—500 нм, которые в некоторых клетках тёмные, а в некоторых — бледные; гранулы разбросаны по всему телу клетки. Железистые клетки наиболее многочисленны в краевой области пластинки (в районе 20 мкм от края), по направлению к центру их численность уменьшается. Волокнистые клетки Клетки соединительнотканного слоя, имеют удивительно ровные края. Отростки волокнистых клеток контактируют с дорсальными поверхностями клеток вентрального слоя и проникают между телами этих клеток, кроме того, они контактируют с дорсальным слоем, другими волокнистыми клетками и липофильными клетками. Впрочем, настоящих клеточных контактов они не образуют. Синцитиальная структура волокнистых клеток не находит чёткого подтверждения. Для волокнистых клеток характерно наличие кластера митохондрий, в который проникают более бледные тельца, палочковидные включения (возможно, эндосимбиотические бактерии), окружённые шероховатым эндоплазматическим ретикулумом, а также крупной конкрементной вакуоли, содержащие остатки водорослей, съеденных организмом. Волокнистые клетки можно визуализировать методом иммунофлуоресценции при помощи антител к предполагаемому кадгерину трихоплакса, а также рецептору P2X[en]. На долю волокнистых клеток приходится 4,4 % всех клеток животного. Дорсальные эпителиальные клетки Подобно клеткам вентрального эпителия, клетки дорсального эпителия соединены межклеточными контактами и имеют реснички и микроворсинки, хотя микроворсинок обычно меньше, чем у клеток вентрального эпителия. Они содержат многочисленные плотные эллиптические гранулы ~0,5 мкм в диаметре. Эти клетки можно визуализировать при помощи антител к MAGUKs и фибриллярному актину. Форма клеток может изменяться от сферической до эллиптической, что соответствует изменениям формы тела животного. Дорсальные эпителиальные клетки составляют 9 % клеток трихоплакса. Кристаллические клетки Кристаллические клетки содержат ромбоидные кристаллы ~ 2 мкм в диаметре, чашевидное ядро, приуроченное к одной стороне клетки, и две центрально расположенные митохондрии, фланкирующие кристалл. Цитоплазма кристаллических клеток лишена включений. Кристаллические клетки лежат под дорсальным эпителием и соседствуют с волокнистыми клетками и липофильными клетками. Они лежат дорсальнее железистых клеток и не имеют входа на поверхность. Точные функции кристаллических клеток неизвестны, но они могут выполнять роль статоцистов или фоторецепторов. Они составляют менее 0,2 % всех клеток в теле трихоплакса.Иногда в старых культурах появляются нежизнеспособные сферические формы трихоплакса. При образовании этих сферических форм дорсальный слой клеток утрачивает контакт с волокнистым синцитием, и слой соединительной ткани наполняется жидкостью. Если при этом вентральный слой клеток отделяется от субстрата, то сферическая форма трихоплакса может некоторое время плавать в культуре как маленький шарик. Выделяют два типа сферических форм: полые, наружная стенка которых представлена ресничным вентральным эпителием, а внутрь полости вдаются волокнистые клетки, причём в полости могут также содержаться ресничные дорсальные клетки; плотные, наружная стенка которых образована дорсальным эпителием, а внутренняя часть плотно набита волокнистыми клетками (кроме того, в ней может находиться компартмент вентральных клеток)[20].
По способу перемещения трихоплакс напоминает амёбу: он медленно скользит по субстрату, постоянно меняя очертания за счёт наличия связей между клетками верхних и нижних слоёв. Постоянные передний и задний концы тела не выражены, поэтому трихоплакс может изменять направление движения, не поворачиваясь. При попытке начать ползти в двух противоположных направлениях одновременно тело трихоплакса может разорваться на две части. Сужение и разрыв перетяжки при делении также происходит за счёт контактов между клетками в верхнем и нижнем слоях[18][21].
Trichoplax adhaerens питается водорослями и другими пищевыми частицами, находящимися на субстрате. Пищеварение внеклеточное, происходит вне тела трихоплакса, между его вентральной поверхностью и субстратом. Во время питания трихоплакс изгибается, приподнимая над субстратом центральную часть, и в образовавшемся замкнутом кармане происходит переваривание пищи; этот процесс иногда называют «временной гаструляцией», причём вентральный эпителий трихоплакса функционально соответствует гастродермису[en] стрекающих. Пищеварительные ферменты, вероятно, выделяются железистыми клетками. Продукты пищеварения поглощаются ресничными клетками, которые могут образовывать эндоцитозные окаймлённые везикулы. По-видимому, маленькое плоское тело делает возможным транспорт путём обычной диффузии, поэтому необходимости в циркуляторной системе у трихоплакса нет[22][16].
Волокнистые клетки содержат конкрементные (пищеварительные) вакуоли, в которых можно обнаружить пищевые частицы на разных стадиях переваривания. Вероятно, отростки волокнистых клеток проникают сквозь верхний слой клеток и там фагоцитируют пищевые частицы[23].
В 2015 году была опубликована статья, в которой сообщалось об интересной детали пищевого поведения трихоплакса. Оказалось, что чем больше водорослей оказалось в пищевом кармане трихоплакса, тем больше липофильных клеток выделяют свой секрет наружу, заставляя клетки водорослей лизироваться. Кроме того, разряжаются только те липофильные клетки, которые находятся на расстоянии 10−20 мкм от пищевой частицы. Механизмы обеспечения такой точной регуляции неизвестны[24].
Имеются сведения о существовании группового поведения у трихоплакса. По некоторым данным, при обитании в больших аквариумах, моделирующих естественные условия, трихоплакс существует в виде скоплений, развитие которых проходит через определённые стадии[25].
Trichoplax adhaerens размножается преимущественно бесполым путём (фрагментацией или почкованием). При фрагментации тело трихоплакса с помощью образующейся перетяжки разделяется на две примерно равные половинки. Процесс фрагментации может занимать несколько часов, причём между расходящимися особями длительное время сохраняется тонкий многоклеточный мостик[26].
При почковании округлые почки (бродяжки) диаметром 40—60 мкм образуются на дорсальной поверхности, они содержат все типы клеток, в том числе вентральные ресничные клетки и соединительную ткань. Формирование почек происходит следующим образом. Зачаток почки формируется во внутреннем слое тела из клеток, которые выселяются внутрь из верхнего и нижнего слоёв, теряя при этом реснички. В дальнейшем эти клетки образуют вентральный эпителий дочерней особи. До отделения почки эти клетки формируют реснички, направленные внутрь полости и, таким образом, меняют полярность на противоположную. По мере роста почка образует выпячивание на дорсальной стороне тела и в конце концов отшнуровывается от неё вместе с частью дорсального слоя клеток. Процесс почкования длится около 24 часов. Отделившись, покрытые ресничками почки уплывают, плавают около недели и оседают на дно. На обращённой к субстрату стороне образуется отверстие, которое постепенно расширяется, так что бродяжка становится чашеобразной, а после распластывается по субстрату и прикрепляется к нему вентральным эпителием. От краевого валика могут отделяться более мелкие почки, образованные клетками дорсального и вентрального слоёв и заполненные волокнистыми клетками. Такие более мелкие почки не способны к плаванию в толще воды и выполнению расселительной функции, они постепенно уплощаются и превращаются в молодых особей[22][27].
Достоверного описания полового процесса у трихоплакса пока нет, однако в лабораторных условиях у него были описаны яйца, а на ультраструктурном уровне — и спермии. Существование полового процесса также подтверждается распределением однонуклеотидных полиморфизмов[13]. Яйцеклетки образуются из клеток нижнего слоя, которые дедифференцируются и погружаются в слой соединительной ткани. Клетки волокнистого синцития играют роль кормящих, часть их отростков вместе с эндосимбиотическими бактериями фагоцитируют яйцеклетки. Половые клетки у трихоплакса образуются нерегулярно; к факторам, запускающим половое размножение, можно отнести увеличение плотности популяции, нехватку пищи и повышение температуры воды до 23 °C и более. И яйцеклетки, и спермии могут формироваться на одной особи. Характер мейоза неизвестен. Яйцеклетка оплодотворяется по достижении размера в 70—120 мкм. После этого яйцо покрывается так называемой защитной «оболочкой оплодотворения» и претерпевает полное равномерное дробление. Зародыш развивается внутри материнского организма, пока последний полностью не разрушится и новый организм не выйдет наружу. В лабораторных условиях зародыши разрушаются на стадии 64 бластомеров по неизвестным причинам (материнская особь тоже гибнет), так что эмбриональное развитие трихоплакса не изучено. Возможной причиной гибели зародыша может быть неконтролируемая репликация ДНК, при которой не происходит перехода клеток из S-фазы в G2-фазу[27][28].
Хромосомный набор Trichoplax adhaerens 2n = 12[18]. Три пары представлены двуплечными мета- или субметацентрическими хромосомами, в то время как три другие пары хромосом имеют меньший размер и, возможно, являются акроцентрическими. Абсолютный размер хромосом не превышает 2—3 мкм[29]. Диплоидные эпителиальные клетки содержат всего 0,08 пг ДНК, что меньше аналогичного значения у любого представителя Metazoa[27].
Секвенированию ядерного генома Trichoplax adhaerens предшествовало секвенирование его митохондриального генома, осуществлённое в 2006 году. Результаты были неожиданными: оказалось, что митохондриальный геном T. adhaerens имеет наибольший размер среди всех Metazoa и содержит более 43 тыс. пар нуклеотидов (в то время как размеры митогеномов других Metazoa обычно лежат в пределах от 15 до 24 тыс. пар нуклеотидов). К отличительным его особенностям следует отнести наличие многочисленных спейсеров между участками транскрибируемой ДНК, нескольких интронов, а также ORF с пока невыясненными функциями (всё это нетипично для других Metazoa), а участки ДНК, кодирующие белки, в среднем на 10 % больше, чем у других животных. Перечисленные особенности митохондриального генома трихоплакса следует считать признаками, унаследованными от общего предка Metazoa, поскольку среди опистоконтов, не относящихся к Metazoa, данные особенности — не редкость. Так, у хоанофлагеллаты Monosiga brevicollis митохондриальный геном содержит примерно 76,5 тыс. пар нуклеотидов, имея большие области, занятые спейсерами, и несколько интронов (а митохондриальный геном аскомицета Podospora anserina[en] состоит из 94—100 тыс. пар нуклеотидов[30])[31][32].
С учётом того, что среди пластинчатых ранее было выявлено наличие значительного генетического разнообразия, нашедшего отражение в выделении нескольких достаточно обособленных клад, исследователи в 2007 году секвенировали митохондриальные геномы ещё у трёх лабораторных образцов, полученных на территории Белиза. Среди них образец под условным обозначением BZ2423 представлял кладу II, BZ10101 — кладу III, а BZ49 — кладу V (сам T. adhaerens принадлежит к кладе I). Анализ этих геномов показал, что они разделяют ранее выявленные у митохондриального генома T. adhaerens особенности: во всех четырёх геномах имеются многочисленные спейсеры, присутствуют несколько ORF с неизвестными функциями, интроны, причём выделен общий набор генов: 12 генов дыхательной цепи, 24 гена тРНК, 2 гена рРНК (rnS и rnL), а гены at8 и atp9 не обнаружены. В то же время наблюдалось значительное различие в размерах генома: у образца BZ49 он содержал 37,2 тыс. пар нуклеотидов, у BZ2423 — 36,7 тыс. пар, а у BZ10101 — 32,7 тыс. пар. Доля кодирующих и структурных последовательностей нуклеотидов варьировала от 55 % у T. adhaerens до 67 % у BZ49. Средняя длина спейсера менялась от 101 пары у BZ10101 и 105 пар у BZ49 до 154 пар у BZ2423 и 209 пар у T. adhaerens[33][31].
Ядерный геном трихоплакса был секвенирован в 2008 году. Он содержит около 98 млн пар нуклеотидов и 11514 белоккодирующих генов, из которых около 87 % обнаруживают значительное сходство с соответствующими генами других животных. Большая часть (около 83 %) из около 7800 семейств генов[en], консервативных у актиний и двусторонне-симметричных животных, имеют гомологи у трихоплакса. Плотность интронов у трихоплакса сопоставима с таковой у позвоночных и актиний. В ядерном геноме Trichoplax сохранились многие древние интроны, утраченные в других линиях животных, и для 82 % интронов человека находятся соответствующие интроны с тем же расположением в генах трихоплакса[12]. Интересно, что некоторые гены трихоплакс приобрёл от своего бактериального грамотрицательного эндосимбионта (риккетсии)[34]. Ядерный геном трихоплакса довольно мал по сравнению с геномами Metazoa, однако это вряд ли можно объяснить вторичным упрощением: об отсутствии вторичного упрощения свидетельствует сохранение древних интронов и порядка генов (синтении)[35].
Несмотря на относительную простоту строения, генетический материал Trichoplax adhaerens содержит множество генов, присущих животным с более высоким уровнем организации[35]. Например, вторичная структура молекулы 16S рРНК у трихоплакса существенно сложнее, чем у любых представителей типа стрекающих (Cnidaria)[36]. Ядерная ДНК трихоплакса содержит гены белков (в особенности, транскрипционных факторов), необходимых для дифференцировки различных типов клеток, развития и функционирования нервной системы, обособления клеток зародышевой линии и других аспектов развития двусторонне-симметричного организма. В ней имеются гены Hox и ParaHox[en] и другие гомеобоксные гены: Forkhead[en], Brachyury и Sox[en]. У трихоплакса также есть гены, кодирующие белки сигнальных путей BMP/TGF[en]- и Wnt, которые чрезвычайно важны для определения осей тела в ходе развития двусторонне-симметричного организма[35].
Кроме того, у трихоплакса обнаружены гены, кодирующие белки базальной пластинки, молекулы, необходимые для межклеточной адгезии и прикрепления клеток к внеклеточному матриксу. Трихоплакс имеет также гены, критичные для работы нервной системы — такие, как гены, кодирующие потенциал-зависимые натриевые[en], калиевые и кальциевые каналы, почти полный набор синаптических каркасных белков[en], белки систем синтеза и транспорта моноаминов, множество ионотропных[en] и G-связанных рецепторов (GPCR), факторы поддержания аксонов и миграции нейронов. Установлено, что у трихоплакса имеются зрительные пигменты — опсины[37]. Имеются доказательства наличия у трихоплакса генов инсулина и инсулиновых рецепторов, а также генов различных регуляторных пептидов и их рецепторов, что свидетельствует о наличии зачатков гормональной регуляции у трихоплакса[13]. У Trichoplax adhaerens обнаружен ген предшественника главного комплекса гистосовместимости (прото-МНС), который необходим для функционирования иммунной системы позвоночных; это рассматривают как аргумент в пользу предположения о раннем возникновении прото-МНС в эволюции животных. У трихоплакса прото-МНС участвует в защитных процессах (противовирусный иммунитет, ответ на стресс, убиквитиново-протеасомный путь расщепления белков)[38].
У трихоплакса обнаружен особый мембранноактивный антимикробный пептид — трихоплаксин, который может быть использован для разработки нового ряда антимикробных препаратов широкого спектра действия[39].
Долгое время вид Trichoplax adhaerens считался единственным представителем типа Placozoa, поскольку в световой микроскоп все пластинчатые из разных частей мира выглядят одинаково. Ряд недавних исследований опроверг это представление. Выяснилось, что среди пластинчатых имеет место значительное генетическое разнообразие, и по состоянию на 2015 год таксон Trichoplax adhaerens в действительности включает 19 видов (пока они именуются гаплотипами), составляющих по меньшей мере 7 хорошо обособленных клад. Описание отдельных видов в случае трихоплакса представляется чрезвычайно сложным из-за однообразной морфологии этих животных[40].
Об экологии и биогеографии трихоплаксов известно очень немного. Пластинчатые обнаруживаются только в спокойных прибрежных водах и избегают глубоководий, а также вод с сильными течениями. Они были найдены и в стоячих водах на глубине до 20 м. Считается, что основными факторами, ограничивающими распространение трихоплаксов, являются температура воды и её солёность. Пластинчатые были обнаружены в тропических и субстропических водах Индийского, Тихого и Атлантического океанов, а именно, вблизи Бермудских островов, в Карибском море, у побережья Восточной Австралии, в Большом Барьерном рифе, около острова Гуам, Гавайев, Японии, в Средиземном море, у островов Палау, Папуа-Новой Гвинеи, в Красном море, вблизи Вьетнама и Восточного Самоа[41]. Члены некоторых клад распространены очень широко, в то время как ареал других клад более ограничен[40].
Пластинчатые предпочитают тёплую воду с температурой до 32 °С, причём иногда они обнаруживались в воде с температурой чуть выше 10 °С. Чаще всего пластинчатые встречаются в местах со спокойными водами на твёрдых субстратах (камнях, коралловых рифах, корнях мангровых деревьев). Разнообразие сред обитания пластинчатых может лежать в основе их большого видового разнообразия. Пластинчатые могут приспосабливаться к мутным водам, а также низкой солёности воды, которая, например, наблюдается в мангровых зарослях[1].
Численность трихоплаксов зависит от времени года. Как правило, пластинчатые наиболее многочисленны в тёплое время года (с июня по октябрь в субтропическом и умеренном поясах Северного полушария)[1].
Наблюдения за связями трихоплаксов с другими живыми организмами показывают, что они часто ассоциированы с сидячими фильтраторами, например, сидячими инфузориями и многощетинковыми червями. Из-за малых размеров и бентосного образа жизни пластинчатые должны часто становиться жертвами хищников, однако пока известен лишь один случай поедания трихоплакса. Описаны случаи, когда потенциальные хищники (например, брюхоногие моллюски или стрекающие) быстро отскакивали после контакта с трихоплаксом, что свидетельствует о возможном существовании химических механизмов защиты у этих животных. Может быть, они опосредуются блестящими шарами, расположенными в верхнем слое клеток[1].
Для сбора пластинчатых из естественных местообитаний существуют два основных метода. Первый из них подразумевает сбор камней, обломков коралловых рифов и других твёрдых объектов с глубины до 5 м. Во втором методе используются обычные предметные стёкла, которые помещаются в пластиковый пакет, открытый с двух сторон, так что через него возможно течение морской воды. Пакеты со стёклами погружаются на глубину от 2 до 20 м. Вскоре стёкла покрываются налётом из водорослей, которые служат кормом для пластинчатых[1]. В лабораторных условиях трихоплаксов кормят одноклеточными водорослями Cryptomonas[en] и Chlorella[41].
Относительно простоты строения Trichoplax нет единого мнения: неизвестно, является ли она первичной, или же она является результатом вторичного упрощения. Существуют даже предположения, что трихоплакс представляет собой прогенетическую личинку какого-то полностью исчезнувшего таксона[27]. Согласно данным анализа митохондриальной ДНК, Placozoa были первой группой, отделившейся вблизи основания эволюционного древа животных. Однако анализ нескольких ядерных генов дал противоречивые результаты. Согласно одним данным, пластинчатых следует рассматривать как базальную группу Metazoa, согласно другим — как вторую ветвь, отделившуюся от древа животных после отделения губок, согласно третьим — как сестринскую группу Bilateria. Анализ 18S рРНК и генов Hox/ParaHox свидетельствует в пользу монофилии группы, включающей таксоны Placozoa, Cnidaria и Bilateria и названной ParaHoxozoa[en]. Недавние палеонтологические находки, проливающие свет на модель питания крупных мягкотелых эдиакарских животных (таких, как Dickinsonia), позволяют заключить, что эти организмы близки к современным пластинчатым, хотя достоверных ископаемых остатков собственно пластинчатых не найдено[42][13].
Многие учёные рассматривали Trichoplax как промежуточное звено между одноклеточными и многоклеточными животными. Под влиянием первых сведений о трихоплаксе О. Бючли выдвинул гипотезу происхождения многоклеточных от плакулы — гипотетической двуслойной колонии. Её нижний слой был обращён к субстрату и выполнял трофическую функцию (будущая энтодерма), а верхний выполнял защитную функцию и дал начало эктодерме. С этой гипотезой хорошо согласуется пищеварительная роль вентрального эпителия Trichoplax. Однако способность волокнистых клеток к трансэпителиальному фагоцитозу больше соответствует модели фагоцителлы, предложенной И. И. Мечниковым[27].
Название типа Placozoa предложил К. Грелль в 1971 году. В противовес этому А. В. Иванов, отдававший предпочтение не модели плакулы, а модели фагоцителлы, предложил выделить тип Phagocytellozoa с единственным родом Trichoplax[4][27]. Несколько позже Иванов (хотя и с замечаниями по поводу «неудачности» названия) признал тип Placozoa, но выделил в системе Metazoa надраздел Phagocytellozoa, включающий единственный тип Placozoa (русский эквивалент названия — Пластинчатые)[43]. Впрочем, позднейшие исследования выявили, что трихоплакс и модель фагоцителлы не имеют между собой ничего общего[44]. Иногда род Trichoplax выделяют в отдельное семейство Trichoplacidae в составе типа Placozoa[45].
Ниже приводится краткий обзор существующих в данный момент гипотез относительно эволюционного положения и происхождения пластинчатых.
Из-за своего простого строения Placozoa часто рассматривались как модельный организм, иллюстрирующий переход от одноклеточных организмов к многоклеточным животным (Metazoa), а потому Placozoa можно считать сестринской группой по отношению ко всем другим метазоям[12]:
MetazoaPlacozoa
Губки (Porifera)
Животные с настоящими тканями (Eumetazoa)
Эта идея, остающаяся в настоящее время чисто умозрительной, получила название функционально-морфологической гипотезы. Она предполагает, что все или большинство животных произошли от галлертоида (от нем. Gallert ‘желе, студень’) — свободноживущего морского пелагического сферического организма. Он состоял из единственного слоя ресничных клеток, поддерживаемого тонким неклеточным разделительным слоем — базальной пластинкой. Внутренняя часть сферы была заполнена способными к сокращениям волокнистыми клетками и желеобразным внеклеточным матриксом[12][46].
Современные Placozoa и другие животные якобы произошли от этой предковой формы, пройдя один из двух путей. Первый путь — впячивание эпителия — привёл к развитию внутренней системы каналов, и такой модифицированный галлертоид с внутренней системой каналов дал начало губкам, стрекающим и гребневикам. Второй путь включал переход от пелагического образа жизни к бентосному. В отличие от морской воды, где пища, потенциальные хищники и половые партнёры могут встретиться со всех сторон, при жизни вблизи субстрата возникает различие между стороной, обращённой к субстрату, и стороной, обращённой к открытому водному пространству. В этих условиях уплощение тела имеет селективное преимущество, и, в самом деле, уплощённое тело присуще многим бентосным животным. Согласно функционально-морфологической модели, пластинчатые и, возможно, некоторые другие группы, известные только из ископаемых остатков, произошли от такой бентосной формы жизни, названной плакулоидом. По этой модели, дорсальный и вентральный слои клеток трихоплакса являются аналогами, но не гомологами эктодермы и энтодермы эуметазоев[12][46].
В случае истинности вышеописанной модели Trichoplax adhaerens оказывался бы древнейшей ветвью многоклеточных организмов и реликтом эдиакарской (или даже пре-эдиакарской) фауны. Из-за отсутствия внеклеточного матрикса и базальной пластинки развитие этих животных, очень успешных в своей экологической нише, было, разумеется, ограничено, что объясняет низкую скорость эволюции пластинчатых и их почти полную неизменность в ходе эволюции[12].
Функционально-морфологическая гипотеза подтверждается анализом митохондриальной ДНК[32][33], однако не находит должного подтверждения в статистическом анализе генома трихоплакса по сравнению с геномами других животных[12].
Согласно гипотезе Epitheliozoa, основанной на чисто морфологических особенностях, Placozoa являются ближайшими родственниками животных с настоящими тканями (Eumetazoa). Образуемый ими таксон (Epitheliozoa), оказывается сестринской группой по отношению к губкам[12]:
MetazoaPorifera
Placozoa
Eumetazoa
Главным подтверждением этой гипотезы является наличие особых клеточных контактов — ленточных десмосом, которые имеются у Placozoa и всех остальных животных, за исключением губок. Кроме того, Trichoplax adhaerens объединяет с эуметазоями наличие железистых клеток на вентральной стороне. В рамках этой гипотезы возможным сценарием происхождения моноцилиарных клеток, похожих на эпителий (эпителиоидных) тканей Trichoplax adhaerens, является редукция воротничка хоаноцитов у группы губок, которая утратила способность к фильтрационному питанию и стала предковой группой пластинчатых. Эпителиоидная ткань трихоплакса впоследствии дала начало эпителию эуметазоев[12].
Согласно модели Epitheliozoa, дорсальный и вентральный слои клеток трихоплакса являются гомологами эктодермы и энтодермы, а слой волокнистых клеток дал начало соединительным тканям других животных. Статистический анализ генома трихоплакса по сравнению с геномами других животных говорит в пользу этой модели[12].
Гипотеза Eumetazoa, базирующаяся в основном на данных молекулярной генетики, рассматривает пластинчатых как крайне упрощённых эуметазоев. Согласно этой гипотезе, трихоплакс произошёл от гораздо более сложно устроенных животных, которые имели мышечную и нервную ткань. Два этих типа тканей, а также базальная пластинка и эпителий были утрачены в ходе последующего радикального упрощения. Было предпринято много попыток определения сестринской группы Placozoa: согласно одним данным, они являются ближайшими родственниками стрекающих, по другим — сестринской группой гребневиков, а иногда их помещают в группу двусторонне-симметричных (Bilateria)[12]:
MetazoaPorifera
Ctenophora
Cnidaria
Placozoa
Bilateria
Главным недостатком этой гипотезы является то, что она практически не учитывает морфологические особенности пластинчатых. Крайняя степень упрощения, которая, согласно этой модели, имеет место у Placozoa, известна только у животных, ведущих паразитический образ жизни и, по-видимому, маловероятна у свободноживущих видов, каким является Trichoplax adhaerens. Статистический анализ генома трихоплакса по сравнению с геномами других животных отвергает гипотезу[12].
Иную картину филогенетических взаимоотношений между основными подразделениями Metazoa дают результаты комбинированного анализа, основанного на совместном использовании морфологических данных, вторичной структуры митохондриальных генов рРНК и нуклеотидных последовательностей из митохондриального и ядерного геномов. На основании этих результатов пластинчатые группируются вместе с губками, гребневиками и стрекающими в кладу Diploblasta, сестринскую к Bilateria[47][48]:
Metazoa DiploblastaPlacozoa
Porifera
Ctenophora
Cnidaria
Bilateria
Объединение упомянутых четырёх типов (Placozoa, Porifera, Ctenophora, Cnidaria) в единый таксон высшего ранга («подцарство Radiata») предложил Т. Кавалье-Смит в своей шестицарственной системе органического мира 1998 года (впрочем, он не настаивал на монофилии данного таксона)[49]. В системе царства Metazoa, предложенной в 2015 году Майклом Руджеро с соавторами и входящей как составная часть в представленную ими макросистему живущих организмов, это царство включает два подцарства: не получившее названия (соответствует группе Diploblasta на приведённой кладограмме) и Bilateria. Данный вариант классификации пригоден и при принятии любой из трёх предыдущих филогенетических гипотез, но тогда указанное безымянное подцарство будет представлять собой уже не кладу, а парафилетическую группу[50][51].
Иногда в старых культурах появляются нежизнеспособные сферические формы трихоплакса. При образовании этих сферических форм дорсальный слой клеток утрачивает контакт с волокнистым синцитием, и слой соединительной ткани наполняется жидкостью. Если при этом вентральный слой клеток отделяется от субстрата, то сферическая форма трихоплакса может некоторое время плавать в культуре как маленький шарик. Выделяют два типа сферических форм: полые, наружная стенка которых представлена ресничным вентральным эпителием, а внутрь полости вдаются волокнистые клетки, причём в полости могут также содержаться ресничные дорсальные клетки; плотные, наружная стенка которых образована дорсальным эпителием, а внутренняя часть плотно набита волокнистыми клетками (кроме того, в ней может находиться компартмент вентральных клеток).
黏丝盘虫(Trichoplax adhaerens)是1883年由德國生物學傢Franz Eilhard Schulze (1840-1921)在奥地利Graz大学的水族馆发现的[1][2]。目前在扁盤動物門中僅確認此一種,一般稱絲盤蟲即指此種。
丝盘虫是已知最简单的多细胞动物之一。丝盘虫为扁平薄片状,直径不超过4毫米,体形经常改变,边缘不规则,无对称性,但有恒定的背腹面。虫体由上下两层细胞构成,无体腔,无消化腔,无神经系统。
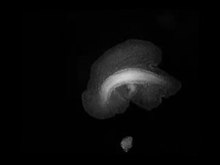 播放媒体
播放媒体
丝盘虫以体表细胞上的鞭毛摆动进行运动。因为缺少神经系统,其躯体各部分的运动方向不能受到协调,有可能躯体两部分同时向两个相反方向运动。其运动方向随机,属于布朗运动。其运动速度受养料浓度影响,养料浓度高的时候,躯体变得扁平,移动变慢,这样有利于扩大吸收面积,有更多时间吸收养料。养料浓度低的时候躯体面积变小,运动加快,以更快的找到新的养料来源。
丝盘虫捕食微小原生动物,以腹面包裹食物,并进行部分体外消化。
丝盘虫的生殖方式为出芽和分裂,虽然也存在有性生殖,但对其胚胎发育知之甚少。
目前已确认的扁盘动物只有黏絲盤蟲(Trichoplax adhaerens)一种。1893年意大利人Franzesco Saverio Monticelli描述了另外一种扁盘动物Treptoplax reptans,据称是在那不勒斯的水域里找到的。但从1896起Treptoplax reptans就被认为是消失了的物种,目前大部分的动物学家都怀疑其存在。
黏丝盘虫(Trichoplax adhaerens)是1883年由德國生物學傢Franz Eilhard Schulze (1840-1921)在奥地利Graz大学的水族馆发现的。目前在扁盤動物門中僅確認此一種,一般稱絲盤蟲即指此種。


センモウヒラムシ (Trichoplax adhaerens) は、単純な風船あるいは煎餅に似た形の海産動物である。軟体性であり、直径は約0.5mm。巨大なアメーバのような見かけだが、多細胞であり、裏表の区別がある。板形動物門[1](または平板動物門、板状動物門 Placozoa)唯一の種(すなわち単型)として分類される。
1883年、オーストリアのシュルツェ(Schulze) によって海水水槽中から発見された。学名である T. adhaerens は、ガラスピペットや顕微鏡のスライドガラスを含む基盤に付着する (adhere) 性質から命名された。これを中生動物とする説がある一方で、同様の動物を観察したStiasnyは、同じ水槽にEleutheria属のクラゲが出現したことに注目し、これを刺胞動物の幼生であるプラヌラの変形したものだと断定した。それ以降は記録がなかったため、種の存在自体が疑問視され、あるいは刺胞動物として決着済みとの文章も一人歩きする状態が続いていた。しかし、1960年代に再発見され、培養に成功したことにより詳細な研究が進んだ。この結果、他の動物群には属さないことが明らかになり、1971年、新たに設けられた平板動物門 (Placozoa) に分類された。
センモウヒラムシは、器官と大部分の組織を欠いている。神経系も存在しないのだが、神経系を備えた種から進化したことを示唆する証拠もある。背と腹の区別があり、3層に分かれた2000-3000個の細胞から構成されている。背側表面は、1本の繊毛を持ち扁平な「扁平上皮細胞」からなる。腹側表面は、1本の繊毛を持ち柱状の「柱状上皮細胞」および、繊毛を持たない「腺細胞」からなる。腺細胞は消化酵素を分泌していると考えられる。これら2層の間には体液で満たされ、「間充織細胞」がある。
かつては無胚葉であると考えられていたが、その後の研究で二胚葉であると考えられるようになってきている。また、近年の分子生物学的な研究では、刺胞動物、有櫛動物(いずれも二胚葉の動物群)との類縁関係が指摘されている。DNA量は約1010ドルトン、小型の原生生物と同程度で、全動物中最も少ない。
水深2-3mの海中にガラス板を沈めておくことで、そこに付着したセンモウヒラムシを容易に採集することができる。体表の繊毛によって移動が可能である。単細胞生物や藻類を食物としている。これらの食物を「腺細胞」から分泌された消化酵素で分解し、体表の細胞で養分を直接吸収している。エサを採るために一時的に体の一部を伸ばすことが観察されている。
全体が2つに分かれる分裂によって無性生殖する。また、背面から多細胞の小塊を作りだす出芽も行なう。さらに、1個体当たり1個か2個の大きな卵細胞を生じることが知られていることから、受精によって有性生殖することができると考えられているが、卵と精子を作り出すための、特別な生殖組織は備えていない。
センモウヒラムシ (Trichoplax adhaerens) は、単純な風船あるいは煎餅に似た形の海産動物である。軟体性であり、直径は約0.5mm。巨大なアメーバのような見かけだが、多細胞であり、裏表の区別がある。板形動物門(または平板動物門、板状動物門 Placozoa)唯一の種(すなわち単型)として分類される。







