 分類 ドメ
分類 ドメアピコンプレックス門(Apicomplexa;またはアピコンプレクサ類)は原生生物界の門の1つ。アピコンプレクサ類は生活環のどこかでアピカルコンプレックス(apical complex、頂端複合構造)という構造を持つという点で特徴づけられる原生生物の大きなグループである。寄生性であり、配偶子の時期を例外として、鞭毛や仮足を持たない。
アピコンプレクサ類の生物はコルポデラ類を除いて全てが寄生性である。宿主は後生動物全般で幅広いが、個々の種は比較的宿主特異性が高いと考えられている。寄生部位も腸管、体腔、組織、血球と様々で、細胞内寄生のものも細胞外寄生のものもある。宿主に対して病原性を示すもの多く知られており、マラリア原虫のような著名なヒトの病原体が含まれている。アピコンプレクサ類によって引き起こされる代表的な疾病には以下のようなものがある。
アピコンプレクサ類は無性生殖と有性生殖からなる複雑な生活環を持っており、一般に次の3つの時期に区分できる。
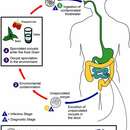
標準的には、宿主はオーシストを摂取することで感染する。まずオーシスト中のスポロゾイトが細胞に侵入してメロゴニーを行い個体数を増やす。生じたメロゾイトは再び周辺の細胞に侵入してメロゴニーを繰り返す(1)。その後、雌雄の生殖母体が分化してガメトゴニーを行い(2)、生じたガメートが受精する(3)とスポロゴニーにより新しいオーシストが生じる(4)。この基本的なパターン以外に様々なバリエーションがあり、また単一宿主のものだけでなく複数の宿主を持つものも多数ある。
アピコンプレクサ類は基本的に単細胞であるが、大きさは数μmから数mmまでと極めて幅がある。細胞膜がアルベオール(alveole、胞室)という小胞に裏打ちされ、やや堅いペリクル(pellicle、外被)を形成している。アルベオールの存在やその他の特徴から、アピコンプレクサ類はアルベオラータというグループに所属させられている。ペリクルは細胞のほぼ全体を覆っているが、飲作用(ピノサイトーシス)を行うミクロポア(micropore、微小孔)の所にはない。
細胞の前端にあるアピカルコンプレックスは、元来はミゾサイトーシスに使われる構造だと考えられているが、大部分の種では宿主細胞との接触や細胞内への侵入に関与していると考えられる。先端は極輪(polar ring)という微小管の帯が取り囲んでおり、多くの原虫にはさらにコノイド(conoid、円錐体)という漏斗状の微小管束がある。開口するロプトリー(rhoptry)とミクロネーム(microneme)という小胞があり、原虫が宿主細胞に侵入するための酵素を分泌している。
もう1つの特徴として、アピコンプレクサ類の細胞にはアピコプラストと呼ばれる細胞小器官があって、3ないし4枚の膜に囲まれていることが挙げられる。その機能は脂質合成などだろうと思われるが、原虫の生存に必須であるらしい。ただしクリプトスポリジウムや(少なくとも一部の)グレガリナ類にはアピコプラストは存在しないと考えられている。アピコプラストと渦鞭毛藻の葉緑体は両方とも紅藻由来だと考えられ、したがって一般的には両分類群の共通祖先から受け継がれたものだと考えられている。しかしアピコプラストは緑藻由来だとする意見もある。
アピコンプレックス門の下位分類は、分子系統解析による再検討が行われている段階である。比較的安定的に受け入れられている分類群は以下の4つであり、これらを綱または亜綱にあてることが多い。
このうち、住血胞子虫類のみ、もしくは住血胞子虫類とピロプラズマ類の2つをコクシジウム類に含めることがある。またピロプラズマ類を住血胞子虫類に含めて扱うこともある。
クリプトスポリジウム(Cryptosporidium)は従来コクシジウム類だと考えられてきたが、最近の分子系統解析によればむしろ特殊化したグレガリナ類であるらしい。実際に培養実験によって、細胞外でグレガリナ類に似た形態をとる時期の存在が判明している。
アピコンプレックス門への所属が議論されるものとして、パーキンサス類とコルポデラ類がある。パーキンサス(ペルキンスス、Perkinsus)は軟体動物に寄生する原虫で、はじめ菌類として分類されていたが、鞭毛虫ステージにアピカルコンプレックス類似の構造が発見されたことでアピコンプレックス門に移された。現在では分子系統解析などにより、アピコンプレクサ類よりも渦鞭毛虫に近縁であると考えられているため、アピコンプレックス門には含めないようになってきている。コルポデラ(Colpodella)は自由遊泳して他の原生生物を吸引消化する鞭毛虫であり、吻(rostrum)がアピカルコンプレックス類似の構造でできている。分子系統解析でもアピコンプレクサ類に近縁であることが示されているが、アピコンプレックス門に含める意見と含めない意見と両論がある。これらの生物はアピコンプレクサ類と渦鞭毛虫の共通祖先に似ているのだろうと考えられている。
アピコンプレックス門は、かつて胞子虫と呼ばれていた寄生性原虫の大部分を含んでいる。もともと胞子虫(Sporozoa Leuckart, 1879)という分類群はグレガリナ類とコクシジウム類からなるものであり、その意味では胞子虫類とアピコンプレクサ類とはほぼ同義である。これ以外に粘液胞子虫類・微胞子虫類・アセトスポラ類をも「胞子虫」として扱っていた期間が長く続いていたが、これはどちらかと言えば便宜上のもので、当時から系統の異なる生物だと考えられていたし、現在ではそれぞれ全く別の生物だと確定している。この経緯については胞子虫の項に詳しい。このような場合、分類学の一般論としては、範囲を狭めて(元に戻して)胞子虫という名を使い続けるのが妥当である。まして胞子虫という名は古典的4分類の1つとして長く親しまれたものであり、アピコンプレックス門という名称が受け入れられるまでには長い論争があった。
獣医学分野で活躍した寄生虫学者Norman D. Levineは、胞子に相当する形態を取らないため胞子虫類でないと疑われていたピロプラズマ類について、アピカルコンプレックスが認められることから胞子虫と近縁と考え、1969年にこれらをまとめてPolannulifera門とすることを提案した。これは翌年Apicomplexa門と改名され、グレガリナ類・コクシジウム類・住血胞子虫類を含む胞子虫綱と、ピロプラズマ綱の2綱が置かれていた。つまり、アピコンプレクサ類は(狭義の)胞子虫類を含みさらに拡大させた概念である。しかし1970年代前半には、ピロプラズマ類は住血胞子虫類と近縁なので独立の綱を与えるべきでなく、したがってアピコンプレクサ類は胞子虫類の同義語で不要と批判された。
ところが1976年に菌類だと考えられていた軟体動物の病原体がアピカルコンプレックスとよく似た構造を持っている事が判明する。Levineはこれにパーキンサスと名付け、アピコンプレックス門を胞子虫綱(Sporozoea、和名では種虫綱と訳し変える)とパーキンサス綱の2綱とした。これが1980年の合意体系で採用され、普及のきっかけとなった。その後の1982年、パーキンサスの構造は渦鞭毛虫によく似ていて、アピカルコンプレックスも本当に相同か疑わしいと批判された。またもやアピコンプレクサ類は胞子虫類の同義語となり不要ということになるが、しかしすでに合意体系で採用されていたこともあり、アピコンプレックス門は普及し続けた。
Levineはその後もアピカルコンプレックスを重視しつづけ、1988年にはアピコンプレックス門にコノイドの有無に着目した2綱(Conoidasida, Aconoidasida)を置くようにし、これによって分類体系から「胞子虫」という名前自体を消してしまった。Levineの死後、分子系統解析によって(狭義の)胞子虫類と(少なくとも一部の)コルポデラ類とが単系統群をなす事が判明しており、アピコンプレックス門という分類群自体は概ね受容されている。一方Conoidasidaという分類群は単系統にはならないため、今後系統を反映した体系に置き換えられることになるだろう。
ウィキメディア・コモンズには、アピコンプレックス門に関するカテゴリがあります。

